
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (20)
Similar a Fitopatologia
Similar a Fitopatologia (20)
Fitopatologia
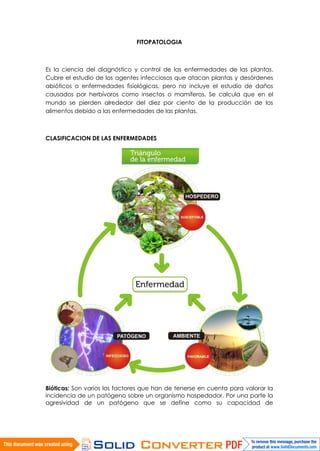
- 1. FITOPATOLOGIA Es la ciencia del diagnóstico y control de las enfermedades de las plantas. Cubre el estudio de los agentes infecciosos que atacan plantas y desórdenes abióticos o enfermedades fisiológicas, pero no incluye el estudio de daños causados por herbívoros como insectos o mamíferos. Se calcula que en el mundo se pierden alrededor del diez por ciento de la producción de los alimentos debido a las enfermedades de las plantas. CLASIFICACION DE LAS ENFERMEDADES Bióticas: Son varios los factores que han de tenerse en cuenta para valorar la incidencia de un patógeno sobre un organismo hospedador. Por una parte la agresividad de un patógeno que se define como su capacidad de
- 2. penetración, de difusión en el hospedador y de crecimiento en los tejidos de éste. Tras la inoculación e incubación del patógeno, el hospedador puede sufrir diversos tipos de daños y aparece la enfermedad. En este caso se dice que el patógeno está dotado de agresividad o virulencia. En caso contrario, se considera desprovisto de ella. Por otro lado los patógenos pueden presentar diferentes grados de dependencia de su hospedador, se diferencia habitualmente dos grupos de parásitos: • Los parásitos estrictos, son altamente dependientes del hospedador. En general no matan las células de las que se nutren. Son específicos o de muy estrecho rango de huéspedes existiendo a veces diferentes cepas o patovares que sólo pueden atacar a especies o incluso razas concretas de plantas. • Los saprófitos tienen un amplio rango de hospedadores, es decir que son polífagos. Matan a la célula por mecanismos enzimáticos antes de alimentarse de su contenido. por lo general son organismos descomponedores de materia muerta que en casos concretos de debilidad en el hospedador pueden colonizar tejidos vivos. El concepto de susceptibilidad es importante a la hora de analizar la incidencia y severidad de un patógeno en un vegetal. Se considera que una planta es susceptible cuando posee la cualidad de convertirse en sede de una infección, es decir, de servir de hospedador a un patógeno. De este modo una planta es susceptible a un patógeno cuando éste es capaz de penetrar, infectar y originar en la planta los síntomas característicos de la enfermedad. Concepto opuesto a la susceptibilidad es la resistencia, propiedad de una planta para oponerse a la penetración y, por tanto, a la infección, no apareciendo síntomas. Esta susceptibilidad puede ser natural, o inducida según dependa de factores genéticos o esté influida por el medio ambiente. Un cualidad intermedia entre susceptibilidad y resistencia es la tolerancia. Un planta es tolerante cuando el patógeno puede penetrar pero no aparece infección ni síntomas. En el desarrollo de una enfermedad pueden diferenciarse diferentes etapas: • Contaminación. Es la llegada del patógeno o del agente infeccioso o inóculo (por ejemplo esporas de hongos) a las proximidades del hospedador. • Penetración del patógeno. Que puede realizarse bien a través del tejido sano de la planta para lo cual el patógeno ha de poseer estructuras (como estiletes de nematodos) o enzimas digestivas que degraden esos tejidos; o a través de heridas o aperturas naturales de la superficie de la planta (como estomas). • Infección. Es el proceso en el que el patógeno toma contacto con las células del hospedador de las que posteriormente va a alimentarse. • Incubación. Es el intervalo de tiempo que transcurre entre la infección y la aparición de síntomas. Depende su duración en gran medida del tipo de patógeno y de los órganos a los que ataque. • Difusión o invasión. Durante esta etapa el patógeno se extiende más allá de los primeros tejidos colonizados. Patógenos muy específicos
- 3. suelen limtarse a áreas concretas de la planta, sin embargo otros patógenos son capaces de colonizar diversos tejidos. • Reproducción del patógeno. Varios grupos de patógenos producen sus estructuras reproductoras en el interior del hospedador, tal es el caso de hongos, virus y bacterias entre otros. • Diseminación o dispersión del patógeno. Las estructuras reproductoras del patógenos alcanzan la superficie del hospedador y son dispersadas en el medio para, bien colonizar nuevas plantas o bien para mantenerse en el exterior como estructuras de resistencia. • Supervivencia. Las estructuras de resistencia como esclerocios en hongos o quistes en nemátodos se mantiene en el medio ambiente siempre que las codiciones ambientales no sean las adecuadas para infectar un nuevo hospedador. Abióticas: Enfermedades NO infecciosas “Causadas por la falta o el exceso de algún factor indispensable para la vida” Ocurren en ausencia de patógenos Pueden afectar plantas en todos los estados fenológicos (semilla poscosecha) y cualquier parte de la planta. Los síntomas varían y dependen del factor involucrado y el grado de desviación del valor normal. CLASIFICACIÓN DE LOS PATÓGENOS DE PLANTAS Hongos La mayoría de los patógenos de plantas son hongos de las divisiones ascomycetes, basidiomycetes u oomycota. Una enfermedad fúngica puede ser descrita como policíclica si el agente causal es capaz de producir esporas y reinfectar plantas durante una temporada de crecimiento, o monocíclica si el agente causal debe esperar una nueva temporada. Esta clasificación aplica a regiones con cuatro estaciones donde los patógenos deben producir estructuras específicas para sobrevivir al invierno. Los hongos fitopatógenos también pueden ser definidos por los tipos de esporas que producen y por el método por el cual penetran en la planta. Una vez que el patógeno ha penetrado produce un haustorio y crece dentro de la planta (alimentación biotrófica), o mata las células de su alrededor y se alimenta del tejido muerto (alimentación necrotrófica). La identificación de los patógenos se realiza basándose en los signos y síntomas de la enfermedad. Signos se refieren a la observación de alguna de las estructuras del patógeno (como esporulación). Síntomas son evidencia secundaria producida por la planta de que un patógeno esta presente (como el marchitarse de las hojas). Las principales enfermedades causadas por hongos son mildius, oidios, royas, carbones, agallas y deformaciones, necrosis, chancros, marchiteces foliares, vasculares, etc., podredumbres radiculares, de flores, de frutos, etc. y micosis post-recolección, etc.
- 4. Procariotas Las bacterias típicamente colonizan espacios intercelulares en distintos órganos o el xilema rodeadas de polisacáridos superficiales, fundamentalmente exopolisacárido (EPS) que aumentan su virulencia. Muchas secretan toxinas que causan daños celulares, entre ellas enzimas que degradan paredes celulares. Algunas, como Agrobacterium tumefaciens transfieren parte de su ADN en un plásmido, el ADN-T, a la célula huésped. Los genes esenciales para la infección (genes de respuesta hipersensible, hrp) aparecen asociados en un cluster de patogenicidad. Muchos de los cuales son similares a los de bacterias patógenas en animales, posiblemente debido a transferencia horizontal y posterior evolución hasta especializarse en la infección de distintos huéspedes. Virus y viroides La patogenia causada por virus es muy característica en cuanto a que el patógeno se incorpora a sí mismo en el metabolismo de la célula hospedadora. Después de infectar una célula vegetal viva el virus se libera de su cubierta proteica e introduce su material genético en el interior del hospedador. El genoma viral se traduce y duplica, ensamblándose numerosas partículas virales nuevas, que utilizan el ácido nucleico y las proteínas recién sintetizadas. El movimiento de los virus desde el lugar de infección a células vecinas se realiza vía simplasto a través de plasmodesmos modificados para permitir el paso de grandes moléculas (como las nucleoproteínas virales), previa inducción de proteínas de movimiento (MPs). Una vez en el floema, la velocidad de movimiento al resto de la planta se incrementa notablemente. Nematodos Los nemátodos infectan siempre el sistema radicular. Son biotróficos, poseen un estilete hueco capaz de atravesar las paredes celulares; muchos modifican el metabolismo de la célula infectada e inducen estructuras especializadas en ella para nutrirse, como las células gigantes resultantes de la fusión de varias células por disolución parcial de sus paredes o por mitosis sin citoquinesis. Algunas formas son ectoparásitos, y se alimentan desde la superficie celular, otros son endoparásitos, e invaden el tejido radicular, donde transcurre gran parte de su ciclo vital, formando quistes. Dentro de los géneros de nematodos fitoparásitos se encuentran Meloidogyne, Xiphinema, Heterodera, Globodera, Pratylenchus, Ditylenchus, Criconemella (Mesocriconema), Helicotylechus, Longidorus, Trichodorus, Paratrichodorus, Belonolaimus, Radopholus, entre otros. Los principales síntomas que ocasiona el ataque de nematodos en plantas son lesiones radicales como agallas o raíces artríticas y la consiguiente manifestación en la parte aérea de la planta en forma de debilidad y problemas en el crecimiento y desarrollo de hojas y frutos debido a la falta de nutrientes. Plantas parásitas
- 5. Son numerosas las especies de vegetales parásitos que viven a expensas de otras plantas bien sobre el pie de planta como el muérdago o en el suelo parasitando las raíces como Cuscuta epithymum. Suele diferenciarse dos tipos de organismo vegetal parásito atendiendo a la presencia o no de clorofila, las plantas holoparásitas son aquella cuya alimentación depende totalmente de su hospedador al ser incapaz de realizar actividad fotosintética, las hemiparásitas poseen una cierta independencia del hospedador aunque por los general no pueden sobrevivir sin él. MICROORGANISMO BENEFICOS PARA LAS PLANTAS Bacterias promotoras del crecimiento vegetal: • Bacterias fijadoras de N2. Son aquellas que se inoculan en la semilla, y cuando ésta germina los exudados radicales en la rizósfera o el rizoplano (zonas de estrecho contacto entre la raíces, suelo y los microorganismos) ó en el interior de raíces para formar nódulos. Asi como en otros sitios de la planta como hojas y tallos, en un suelo pobre o restringido de nitrógeno combinado, en esa condición sucede la actividad de fijar N2 del aire. Ejemplos de este tipo son: Azospirillium spp, Azotobacter spp en gramíneas como el maíz y el trigo, Burkholderia vietnamiensis , aunque la cianobacterias mas populares para la inoculación de arroz en Asía y en algunas regiones de México son: Anabena, Nostuc una clase de bacteria común en mantos de agua del mundo y que desde hace mucho tiempo se reporta como un asociado que proporciona beneficio en el ahorro de fertilizante nitrogenado en la producción de arroz. Pero sin duda el grupo bacteriano mejor conocido y famoso del mundo es el que fijan nitrógeno molécular en asociación con leguminosas es Rhizobium y sus asociados considerado uno de los más importantes pues desde su descubrimiento en Europa, se emplea de manera comercial para la producción agrícola en prácticamente cualquier suelo del planta, el cuadro 1 muestra los tipos de inoculantes para leguminosas a base de este grupo que existen en el mercado internacional y que en centros de investigación como el IIQB-UMSNH se pueden conseguir ejemplos: Rhizobium etli para frijol, Bradyrhizobium japonicum para soya al igual que otras leguminosas, en donde se señalan las diversas posibilidades de aplicación de del género Rhizobium. En general como todo lo biológico se requiere conocer lo básico en el manejo para obtener el máximo provecho en términos de ahorro en la aplicación del fertilizante nitrogenado, rendimiento rentable y conservación del capacidad productiva del suelo, en especial en un agricultura orgánica y sustentable. Mientras Frankia spp un actinomiceto (tipo de bacteria que por su forma de crecimiento se recomienda inocular en especies forestales para programas de reforestación e incluso para fitobiorremediación de suelos contaminados con hidrocarburos y otros agentes. en especies forestales Bacterias que solubilizan de fosfatos.
- 6. Las cuales se emplean cuando las semillas se siembran en suelos ácidos u alcalinos de pH 5 a pH 8, lo que provoca, la precipitación de los fosfatos, así como su unión, en consecuencia las raíces vegetales sufren de stress nutrimental por falta de fosfatos (10). Si el suelo carece de suficiente fosfato disponible o móvil entonces se recomienda su inoculación a la siembra de la semilla de esta forma se resuelve la disponibilidad de fosfatos soluble del cultivo vegetal y se evita un bajo o pobre rendimiento (8,9), un ejemplo de esta clase de bacterias es el género Bacillus spp, Arthrobacter spp, Azotobacter spp y otros, lo que obviamente incluye al grupo mas conocido para resolver el problema de movilidad de fosfatos en el suelo las micorrizas de tipo ecto como endotrofico , que serán tratadas en otro articulo. MECANISMOS DE CONTROL BIOLÓGICO DE ENFERMEDADES POR LAS PLANTAS POR BACTERIAS A) Antibiosis. La bacteria coloniza las raíces en crecimiento y libera moléculas antibióticas alrededor de la raíz, perjudicando así los patógenos próximos a la raíz (indicado por las estrellas). (B) La Resistencia Sistémica Inducida (ISR). Muchos productos bacterianos inducen el sistema de señalización, lo cual puede resultar en la protección de toda la planta contra las enfermedades causadas por organismos diferentes. (C) Competencia. Las bacterias de control biológico actúan compitiendo por los nutrientes y los nichos que ocupan en la raíz. Respuesta de las plantas al ataque de patógenos Cuando un patógeno ataca una planta, en el tejido vegetal se activan unos genes que actúan degradando la pared celular de esos microorganismos ó destruyendo las células infectadas de la planta. Por el contrario, si se trata de un daño mecánico debido al ataque de insectos, se activan genes inhibidores de proteasas, lo cual causa toxicidad en los insectos. PLANTAS TRANSGÉNICAS Plantas resistentes a enfermedades producidas por virus, bacterias o insectos. Plantas con capacidad de producir antibióticos, toxinas y otras sustancias que atacan a los microorganismos. Desarrollo de plantas que den frutos de maduración más lenta. Mejora de la calidad de las semillas
- 7. Aplicaciones farmacéuticas. Plantas con mayor producción Mejoramiento en la calidad Nutritiva La Ingeniería genética permite el acceso y manipulación directa de los genes rompiendo las barreras impuestas por la divergencia genética. Esta tecnología nos permite no sólo introducir en una planta genes procedentes de otras especies vegetales sino también de animales y microorganismos. De esta manera se obtienen plantas transgénicas, es decir, portadoras de un gen ajeno o exógeno que se denomina transgén. Para llegar al nivel actual de desarrollo de esta rama de la ingeniería genética vegetal ha sido necesaria la aportación de los importantes avances en el conocimiento de la Biología molecular de los ácidos nucléicos y el desarrollo de la técnica del cultivo de tejidos vegetales in vitro. Las plantas transgénicas tienen en potencia múltiples aplicaciones y a continuación se nombran algunas, muchas de ellas con una importante implantación en el mercado agrícola a finales del siglo XX: -Incremento de la productividad al proteger los cultivos -Regeneración de suelos contaminados por metales pesados con plantas transgénicas tolerantes a concentraciones elevadas de estos elementos. -Producción de medicamentos. En 1997 se investigaba la producción de anticuerpos monoclonales, vacunas y otras proteínas terapéuticas en plantas transgénicas de maíz y soja. -Retraso de la maduración de los frutos para conseguir dilatar el tiempo de almacenamiento. Procedimientos para la obtención de plantas transgénicas Se emplean principalmente tres métodos para introducir genes ajenos en una planta. Todos estos métodos obtuvieron por primera vez, con más o menos éxito, plantas transgénicas en la década de los ochenta y muchas de ellas se comercializaron en los noventa. El primer método que se ideó se basa en el mecanismo natural de infección de la bacteria del suelo Agrobacterium tumefaciens que introduce un gen de su plásmido en las células de la planta infectada. Recordemos que un plásmido es un fragmento de ADN circular y extracromosómico que suele contener información no vital para la bacteria y cuyo tamaño es del orden del 1 al 3% del cromosoma bacteriano (fig. 1 y 3). Este gen se integra en el genoma de la planta provocándole un tumor o agalla. Se aplicó con éxito por primera vez en 1984 en el tabaco y el girasol. Las gramíneas y en general todas las monocotiledóneas presentan gran resistencia a Agrobacterium por lo cual este método es bastante inviable en un extenso grupo de plantas de gran importancia económica.
- 8. Otro método empleado para transformar genéticamente plantas es el uso de protoplastos, que son células vegetales a las que se les ha liberado de la pared celular. De esta manera queda eliminada la barrera principal para la introducción de genes foráneos. Mediante esta técnica se consiguió por primera vez cereales transgénicos en 1988. En el año 1987 se inventa el método del microcañón o cañón de partículas que consiste en bombardear tejidos de la planta con micropartículas metálicas cubiertas del fragmento de ADN que interesa se integre en el ADN de la planta. Es el procedimiento que más éxitos ha conseguido y el que promete más avances. TRANSFERENCIA GENÉTICA CON AGROBACTERIUM TUMEFACIENS En 1970 se planteó la hipótesis de que la enfermedad de las plantas denominada agalla del cuello podría ser producida por la transferencia de material genético entre una bacteria, Agrobacterium tumefaciens, y las células vegetales. La agalla del cuello se caracteriza por la formación de voluminosas agallas, sobretodo en el cuello del tallo (zona de contacto entre el tallo y la raíz), también en las raíces y el tallo de numerosas plantas de interés agronómico. La enfermedad es de naturaleza tumoral y ya se había demostrado, a finales de los años sesenta, que las células afectadas contienen unas sustancias, las opinas (sustancias nitrocarbonadas), que no se encuentran en las células normales. También se demostró que existen varias clases de tumores en función de la concentración de opinas y que es el material genético de la bacteria el que determina este carácter ya que estas observaciones se realizaron en tejidos cultivados in vitro, es decir, en ausencia de bacterias . Se concluyó que las células tumorales habían adquirido la propiedad de sintetizar opinas durante la interacción con la bacteria. También se concluyó que la naturaleza de las opinas depende de la cepa bacteriana y también que cada cepa degrada específicamente sus propias opinas. Quedaba demostrada la hipótesis de la transferencia de información entre la bacteria y la célula vegetal. En resumen la bacteria no es patógena per se porque no segrega ninguna toxina que disuelva las paredes celulares como hacen otras bacterias patógenas. Sus efectos se deben a la transferencia de un segmento de ADN, el ADN-T, cuya expresión en las células vegetales es la causa de la enfermedad. La supresión en el plásmido del segmento transferido hace que la bacteria sea inofensiva sin que ello se la prive de la capacidad de transferir ADN a una célula vegetal. Por tanto se puede plantear su sustitución por un fragmento de ADN extraño. El segmento ADN-T está delimitado en ambos extremos por unas secuencias determinadas de nucleótidos que actúan a modo de señales. La señal "promotor" al principio y la "terminador" al final. La región transferida y que se integra en el genoma de la planta es la comprendida entre estas dos señales. En teoría era posible transferir cualquier gen extraño colocado entre estas dos secuencias. En 1983 se introdujo un gen bacteriano que confería resistencia al antibiótico cloramfenicol. Se escogió este gen sólo porque es fácil poner de
- 9. manifiesto su expresión: las células que han integrado el gen sintetizan el enzima cloramfenicol transacetilasa que gobierna la síntesis del antibiótico. El gen empleado se expresa en la bacteria Escherichia coli. Para que un gen pueda expresarse el enzima ARN polimerasa debe reconocer el "promotor" y el "terminador". La ARN polimerasa del tabaco (una planta muy empleada en estos experimentos de transferencia de genes) no reconoce los promotores y terminadores de E. coli y por consiguiente no transcribe este gen. Para solucionar el problema se fabricó un gen compuesto o quimérico a partir del gen de la resistencia al cloramfenicol de E. coli, un promotor y terminador procedentes del segmento ADN-T de Agrobacterium tumefaciens. El gen quimérico se reincorporó en un plásmido Ti Gen quimérico en el plásmido Ti de Agrobacterium tumefaciens. De esta manera el gen quimérico funcionó al poder ser detectada la actividad de la cloramfenicol transcetilasa en tejidos tumorales. Aún quedaba una dificultad a salvar: la regeneración de una planta entera a partir de células transformadas. Como las células transformadas eran tumorales eran incapaces de esta regeneración y el siguiente paso consistió en eliminar los genes tumorales del segmento ADN-T. De esta manera se pudo regenerar plantas enteras transgénicas que eran fértiles y con las que se pudo estudiar la transmisión de caracteres a su descendencia. Además si se escogen los promotores adecuados, es posible expresar genes en órganos específicos, como raíces, semillas y tubérculos. El gen de la resistencia a antibióticos no tiene interés agronómico por lo que había que identificar, aislar y clonar los genes que pudiesen mejorar las plantas cultivadas. En el caso de caracteres con base genética compleja (donde intervienen numerosos genes), como la resistencia de una planta al frío, es mucho más difícil la manipulación genética que con los caracteres que se expresan como consecuencia de la actividad de un enzima.
- 10. El sueño de obtener plantas resistentes a los insectos fitófagos se ha hecho realidad con la obtención de plantas transgénicas portadoras de un gen bioinsecticida. Bacillus thruringiensis es una bacteria grampositiva del suelo que en los estadios de esporulación produce unos cristales de proteínas de propiedades insecticidas. Berliner en 1909 aisló la bacteria de los cadáveres del gusano de la harina (Ephestia kuehniella) procedente de Turingia. Al creerse que la bacteria era el causante de la muerte del insecto, sugirió la idea de recurrir a B. thuringiensis para luchar contra la plaga de insectos. Los primeros preparados comerciales aparecieron en 1938. Era práctica habitual en los agricultores tirar a voleo esporas de B. thuringiensis sobre los cultivos pero se presentaba el inconveniente de tener que realizar la práctica con una frecuencia mucho mayor que con los insecticidas químicos. A estas proteínas se las denominó cry (del inglés crystal) por su capacidad de formar cristales o ð-endotoxinas por su acumulación en el interior de las bacterias y su carácter tóxico. Las proteínas cry provocan la lisis de las células intestinales de los insectos. Estos bioinsecticidas se caracterizan por su especificidad, pues sólo son tóxicos en escarabajos, moscas y mariposas (grupos de insectos causantes de la mayoría de las plagas), y porque son prácticamente inocuas en humanos. E. Schnepf y H. Whiteley aislaron en 1981 el primer gen que codifica una proteína insecticida. Se acababa de sentar las bases para que M.D. Chilton en 1983 obtuviera las primeras plantas transgénicas de tabaco utilizando Agrobacterium tumefaciens. Le siguieron otros experimentos en diversos laboratorios de Europa y América con el tomate y la patata. Estos experimentos sirvieron para demostrar que la expresión de proteínas insecticidas en plantas era posible y proporcionaba un método eficaz de lucha contra los insectos Obtención de plantas transgénicas resistentes a los insectos mediante Agrobacterium tumefaciens.
- 11. Todas estas investigaciones culminaron en 1996 con la entrada en el mercado de plantas transgénicas (algodón, patata y maíz) resistentes a insectos. A todas estas plantas transformadas se las denomina Plantas Bt (de Bacillus thuringiensis). En 1997 el 25% de los cultivos transgénicos comercializados portaban genes cry. El problema de la aparición de insectos resistentes a estas plantas se prevé solucionarlo con la implantación de distintas proteínas insecticidas en una misma planta transgénica o en plantas transgénicas plantadas en años alternativos.