Descargar para leer sin conexión




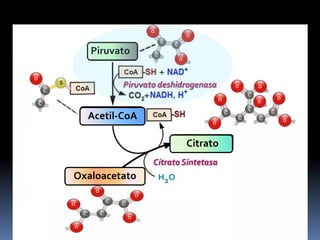
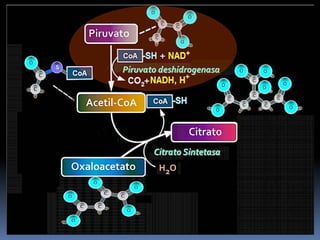




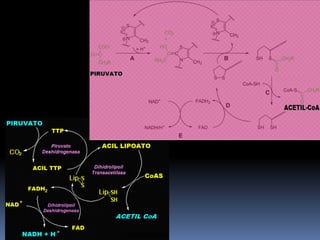






























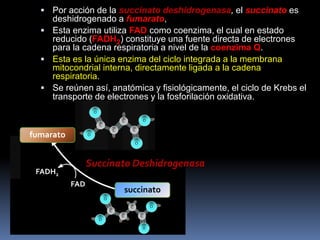




El documento describe las reacciones enzimáticas de la conversión del piruvato a acetil-CoA a través de la piruvato deshidrogenasa, y las primeras etapas del ciclo de Krebs, incluyendo la formación de citrato a partir de la condensación de acetil-CoA y oxalacetato catalizada por la citrato sintetasa, y la conversión de citrato a isocitrato por la aconitasa.



































































