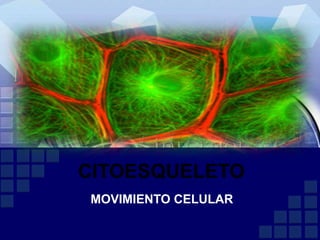
El documento describe la función del citoesqueleto en el movimiento celular y su papel en la mitosis. El citoesqueleto está compuesto de microfilamentos de actina, microtúbulos y filamentos intermedios que dan forma y soporte a la célula, facilitan el transporte intracelular y la división celular. Los microfilamentos de actina se involucran en la motilidad celular mientras que los microtúbulos guían el movimiento de organelas y cromosomas durante la mitosis. Diferentes proteínas regulan la formación y organización de estas e





































































































![Citoesqueleto y movimiento_celular_yboa[1]](https://cdn.slidesharecdn.com/ss_thumbnails/citoesqueletoymovimientocelularyboa1-140622103639-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)






































