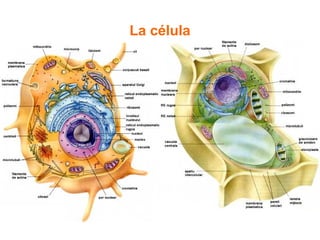
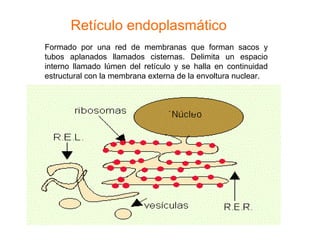
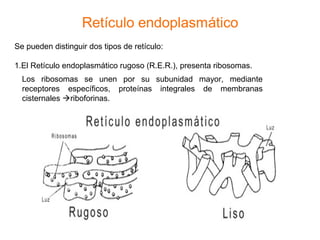
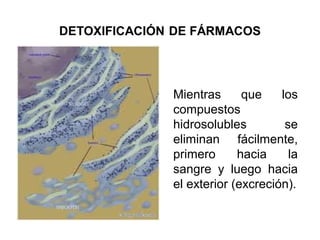
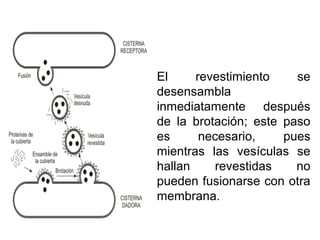
El documento describe los diferentes compartimentos intracelulares, centrándose en el retículo endoplasmático rugoso y liso, y sus funciones en la síntesis de macromoléculas, detoxificación y metabolismo de lípidos y carbohidratos. También se explica el papel del aparato de Golgi en la modificación y exportación de proteínas y lípidos. Finalmente, se abordan temas de transporte vesicular y la función de las vacuolas en la regulación del volumen celular.