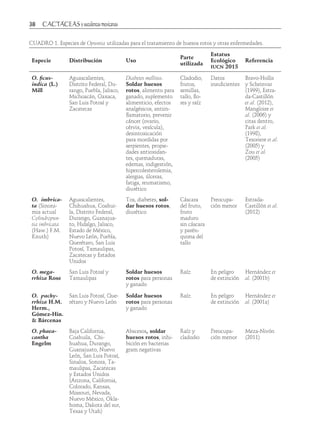
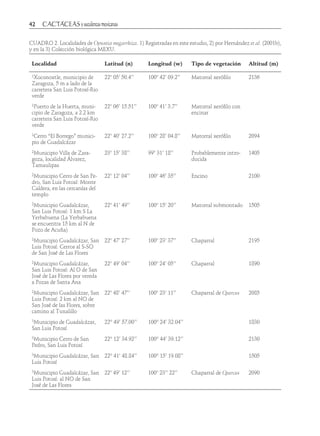
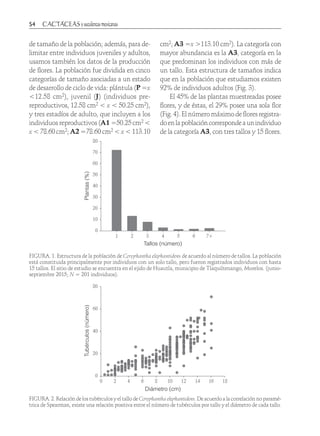
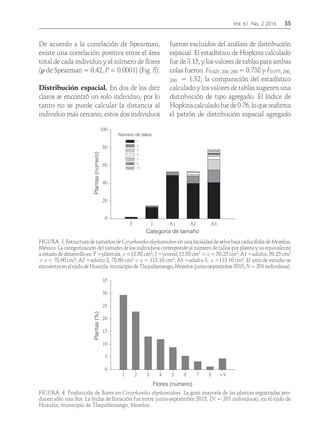
Descargar para leer sin conexión



















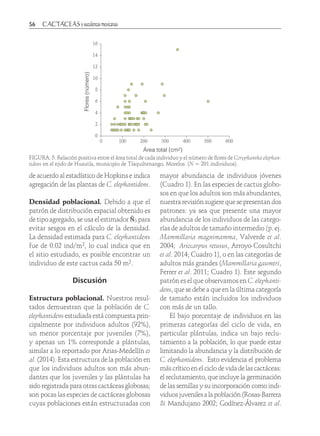

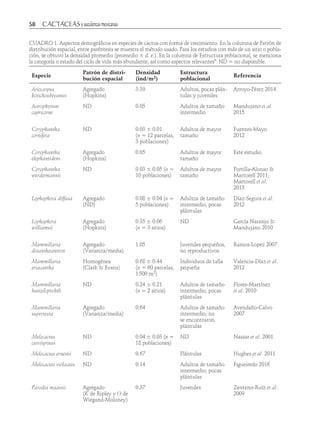







Este estudio documentó el conocimiento tradicional y uso medicinal de Opuntia megarrhiza en San Luis Potosí, México. Se describieron y compararon las estructuras poblacionales de tres poblaciones de O. megarrhiza. Dos poblaciones presentaron reclutamiento de nuevos individuos, aunque pocos alcanzaron la edad adulta. Una tercera población en una zona de pastoreo contenía individuos en casi todas las categorías de tamaño, pero con baja frecuencia. Aún se usa O. megarrhiza para curar

























































