Descargar para leer sin conexión




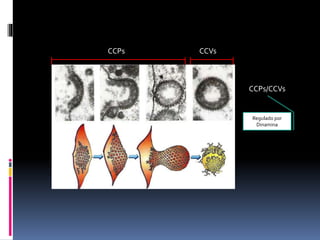


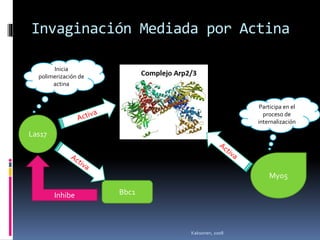
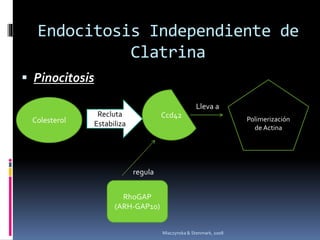
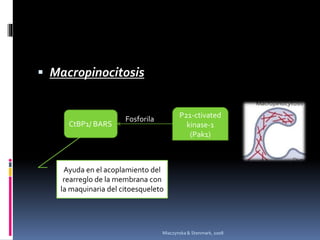

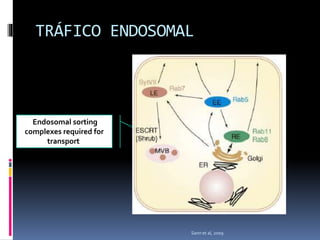
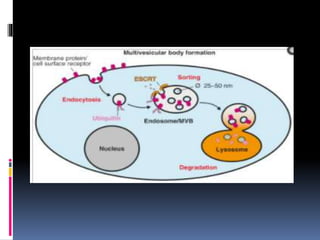





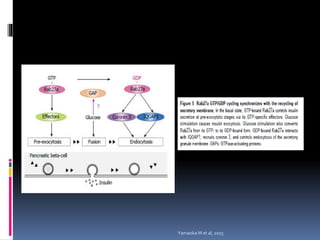

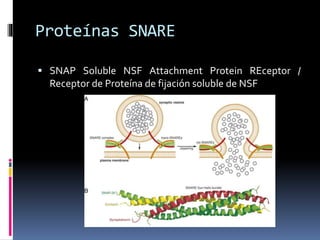

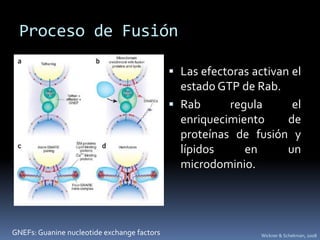









El documento revisa los procesos de endocitosis y exocitosis, destacando sus funciones y mecanismos, incluyendo sus modalidades dependientes de clatrina y apocrina. También se describen las proteínas involucradas en la fusión de membranas y los diferentes tipos de secreción celular. Se menciona la importancia de estos procesos en la comunicación y nutrición celular.















































































