Descargado 17 veces
















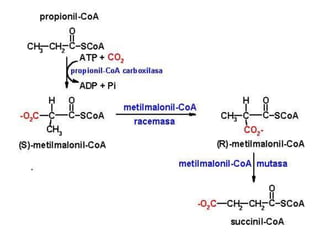




El documento describe la ruta metabólica de la beta oxidación para la oxidación de ácidos grasos. La beta oxidación consiste en una secuencia de cuatro reacciones que separan fragmentos de dos carbonos del extremo carboxilo del ácido graso, resultando en la formación de una acil-CoA acortada. Este proceso se repite hasta que el ácido graso se oxida completamente en acetil-CoA, liberando una gran cantidad de energía en forma de ATP.













































































