Descargado 11 veces





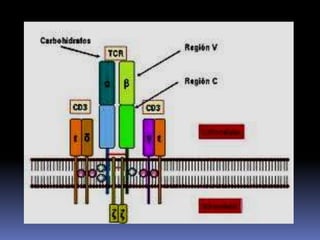

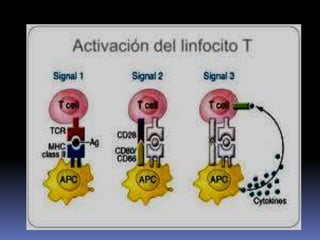


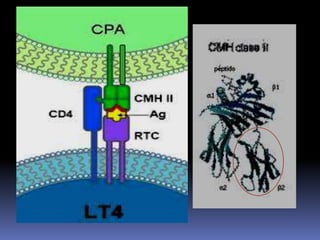




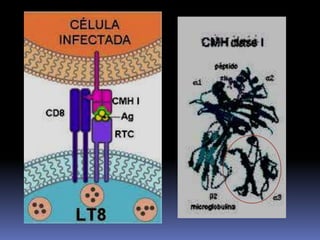


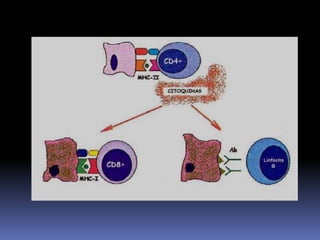

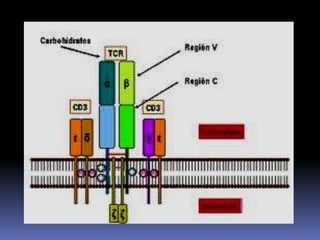

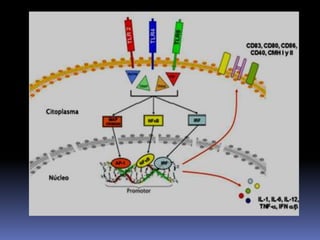




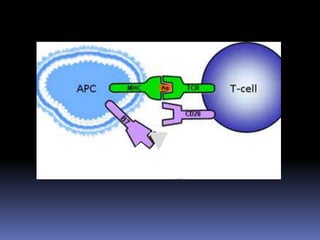


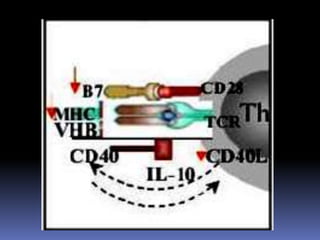





Este documento describe los procesos de activación de los linfocitos T. Explica que los antígenos son captados por células dendríticas y transportados a los ganglios linfáticos, donde son presentados a los linfocitos T junto con moléculas coestimuladoras. Esto causa la activación de los linfocitos T a través de señales mediadas por el receptor del antígeno y moléculas coestimuladoras. Los linfocitos T activados se diferencian en células efectivas que ayudan a eliminar













































































