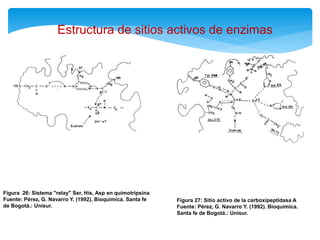
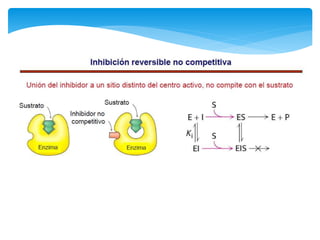
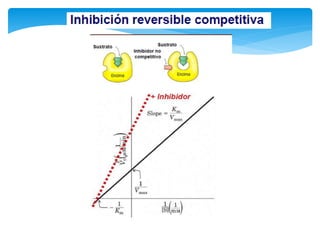
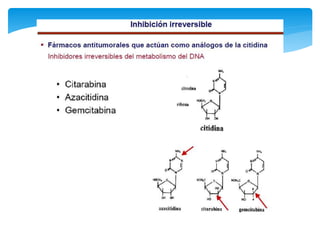
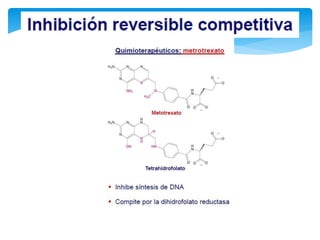
Este documento describe los diferentes parámetros que influyen en la velocidad de reacción enzimática, incluyendo la cantidad de enzima, la concentración de sustrato, la temperatura, el pH, y la presencia de inhibidores. Explica los tres tipos principales de inhibición enzimática: competitiva, no competitiva e irreversible, y cómo cada uno afecta la curva de Michaelis-Menten. También describe ejemplos específicos de enzimas y sus sitios activos.



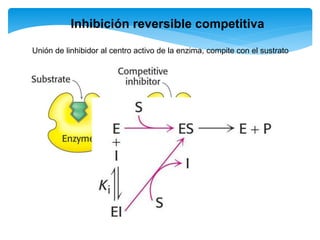
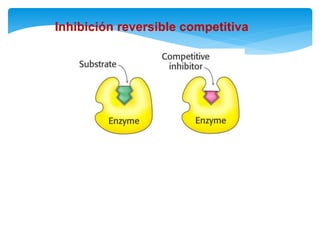

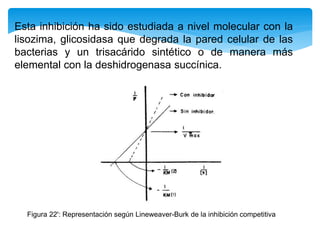
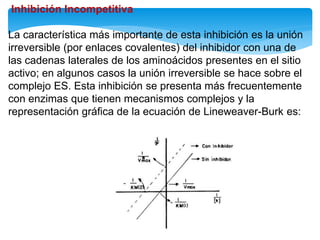
![Inhibición competitiva
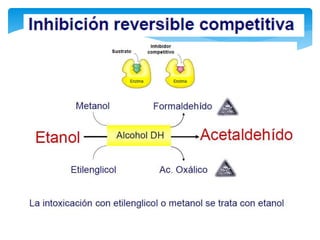
En este tipo de inactivación, el inhibidor (I) posee una estructura química similar a la
del sustrato: esto le permite fijarse reversiblemente en el sitio activo de la enzima
excluyendo el sustrato que previamente se haya unido. La enzima no modifica el
inhibidor porque éste no posee enlaces susceptibles al ataque catalítico, que si están
presentes en el sustrato. La situación puede representarse con las reacciones:
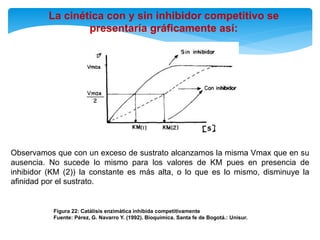
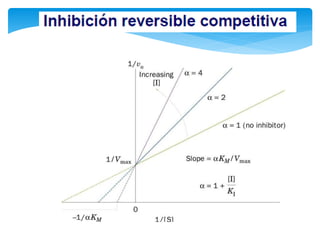
De acuerdo con esto podemos deducir que la magnitud de la inhibición dependerá
de la [I] y que un exceso de S elimina la inhibición pues se usará toda la enzima
libre y el equilibrio se desplazará hacia la formación de ES, y de allí a la formación
de P.](https://image.slidesharecdn.com/inhiidoresenzimaticos-151009141022-lva1-app6891/85/Inhiidores-enzimaticos-13-320.jpg)