
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Similar a Seminario 1 Transporte en Membrana 2021 10.pdf
Similar a Seminario 1 Transporte en Membrana 2021 10.pdf (20)
Más de ANGIEANDREAALFAROAND
Más de ANGIEANDREAALFAROAND (7)
Último
Último (20)
Seminario 1 Transporte en Membrana 2021 10.pdf
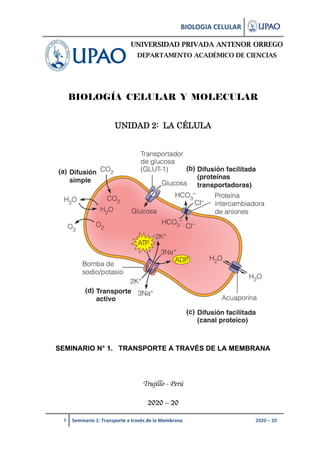
- 1. BIOLOGIA CELULAR 1 Seminario 1: Transporte a través de la Membrana 2020 – 20 BIOLOGÍA CELULAR Y MOLECULAR UNIDAD 2: LA CÉLULA SEMINARIO N° 1. TRANSPORTE A TRAVÉS DE LA MEMBRANA Trujillo - Perú 2020 – 20 UNIVERSIDAD PRIVADA ANTENOR ORREGO DEPARTAMENTO ACADÉMICO DE CIENCIAS
- 2. BIOLOGIA CELULAR 2 Seminario 1: Transporte a través de la Membrana 2020 – 20 PAUTAS PARA EL DESARROLLO DE LOS SEMINARIOS El seminario es una técnica de enseñanza, basada en el trabajo en grupo e intercambio oral de información, utilizada para trabajar y profundizar desde el debate y análisis colectivo en un tema predeterminado. • Desarrollar habilidades de interpretación de información y síntesis de la misma. • Desarrollar la capacidad de comunicación científica, y sustentación oral de un tema a exponer. 1. Los seminarios son de realización obligatoria. 2. Todo Seminario concluye con un resumen, una evaluación de la exposición realizada, así como una evaluación escrita de los temas tratados. 3. El docente moderador: − Presentará e introducirá el tema a exponer − Informará sobre la forma de participación − Ordenará las intervenciones de los participantes, además fomentará la discusión con la participación activa de los estudiantes y aclarará sus dudas. − Velará por la disciplina del grupo. 4. Los alumnos − Deberán tener su videocámara encendida y apagarán sus micrófonos durante la sesión. − Tendrán un buen comportamiento durante la sesión de clase − Deberán leer la información proporcionada por el docente. − Participarán activamente, escuchando a los expositores, para intervenir cuando llega el momento, mediante aportes complementarios, preguntas, aclaraciones o rectificaciones. Los resultados del seminario dependen de sus aportes, pues debe haber leído los temas a tratar y debe tener algo que aportar. − Construirán esquemas o gráficos de los temas considerados en el seminario. Estos esquemas o gráficos deben estar bien representados; pues le servirá para explicar los ítems a desarrollar. Muchos estudiantes creen que preparar el seminario es una tarea sencilla que pueden comenzar y terminar en pocos días. Éste es el error más grande que se puede cometer. Si se deja la preparación para último momento es casi seguro que no podrá desarrollar adecuadamente su presentación. Usted estará más tranquilo si la presentación está lista varios días antes de la fecha asignada, lo que le permitirá a su vez aclarar algunos puntos que tenga duda, con el docente. Puede buscar información adicional, según la bibliografía considerada en el sílabo. Mientras más conozca sobre el tema, más completa será la presentación, más confianza tendrás en tu preparación y mejor contestarás las preguntas. Es muy conveniente que los alumnos tengan a la mano los elementos de información y consulta necesarios (fichas bibliográficas, libros o revistas) para fundamentar la respuesta a la pregunta formulada por el docente moderador. OBJETIVOS NORMAS PARA DESARROLLAR UN SEMINARIO COMO PREPARARSE ADECUADAMENTE
- 3. BIOLOGIA CELULAR 3 Seminario 1: Transporte a través de la Membrana 2020 – 20 SEMINARIO N° 1 TRANSPORTE A TRAVÉS DE LA MEMBRANA Fecha: 26 de octubre al 07 de Noviembre de 2020 1. COMPETENCIAS ESEPECÍFICAS 1. Describe las características básicas de los principales tipos de lípidos encontrados en las membranas celulares y el papel que cumplen cada una de ellas. 2. Describe las características de las tres clases de proteínas de membrana (integrales, periféricas y fijadas a la bicapa) y, como se diferencian unas de otras. 3. Clasifica los diferentes tipos de transporte a través de las membranas celulares de acuerdo a las características funcionales que los distinguen, especialmente las diferencias entre los mecanismos activos y pasivos. 4. Diferenciar los términos gradiente de concentración y gradiente químico y detalla las concentraciones iónicas normales en los diferentes compartimientos corporales (medios intracelular y extracelular). 5. Define difusión simple y explica los factores que pueden afectar el transporte de iones o moléculas por este mecanismo. 6. Define difusión facilitada y describe las características de las proteínas transportadoras 7. Explica el concepto de canal iónico y clasifica los diferentes tipos de canales iónicos en base a su mecanismo de activación, y sus consecuencias sobre la permeabilidad selectiva de la membrana. 8. Describe el papel de las permeasas en la difusión facilitada 9. Explica el papel de los fenómenos osmóticos en los movimientos de agua a través de las membranas celulares y describe las modificaciones inducidas en el volumen celular al ponerse en contacto con soluciones isotónicas, hipertónicas e hipotónicas. 10. Define transporte activo y, establece las diferencias entre los distintos tipos de bombas del transporte activo primario. 11. Describe las bombas de tipo P y explica la actividad de la Na+ /K+ ATPasa en la homeostasis celular. 12. Explica las características estructurales de las bombas de tipo V, F y ABC y, menciona ejemplos de cada uno de ellos. 13. Describe los mecanismos de transporte activo secundario: cotransportadores e intercambiadores y menciona ejemplos de cada uno de ellos. 14. Describe los mecanismos implicados en el transporte epitelial o transcelular, y explicar las consecuencias funcionales de la distribución polarizada de diferentes proteínas de transporte en el movimiento de sustancias a través de las células epiteliales.
- 4. BIOLOGIA CELULAR 4 Seminario 1: Transporte a través de la Membrana 2020 – 20 2. TEMAS 1. Estructura de la membrana celular 1.1. Composición lipídica y organización espacial en la membrana. - Lípidos: Glicerofosfolípidos, esfingofosfolípidos, glucolípidos y colesterol - Influencia de la composición lipídica en las propiedades de la membrana - Asimetría de los lípidos de la membrana 1.2 El componente proteico de las membranas - Proteínas interactúan con las membranas en tres diferentes vías: Extrínsecas o periféricas, Intrínsecas o integrales y ancladas a la bicapa 2. Mecanismos de transporte de moléculas a través de las membranas: concepto y diferencias entre transporte activo y pasivo. 3. Concepto de gradiente de concentración, gradiente de voltaje o potencial eléctrico y gradiente electroquímico. 4. Transporte pasivo 4.1. Difusión simple - Concepto y ejemplos de sustancias que utilizan este mecanismo de transporte. - Factores que influyen en la velocidad de transporte a través de membrana. - Difusión simple de iones a través de proteínas canal en la membrana. - Difusión simple de moléculas de agua (ósmosis) a través de aquaporina (canales de agua) en la membrana 4.2. Difusión facilitada a. Difusión mediada por acarreador (definición ejemplos) 4.3. Transporte activo: 4.3.1. Transporte activo Primario: a. Bomba clase P (características, importancia). Bomba de Na+ /K+ , bomba de H+ /K+ , bomba de Ca2+ b. Bomba clase F (características, importancia, ejemplos) c. Bomba clase V (características, importancia, ejemplos) Bomba de H+ d. Superfamilia ABC (características importancia) CFTR MDR 4.2.2. Transporte Activo Secundario: a. Contratransporte. (características, importancia y ejemplos) b. Cotransporte. (características, importancia y ejemplos) 5. Transporte transepitelial
- 5. BIOLOGIA CELULAR 5 Seminario 1: Transporte a través de la Membrana 2020 – 20 TRANSPORTE A TRAVÉS DE LA MEMBRANAS CELULARES 1. Cuál es la composición química de las membranas celulares ? Qué lípidos se encuentran en las membranas celulares ? El análisis bioquímico y ultraestructural de las membranas biológicas revela que están compuestos por lípidos, proteínas y una pequeña fracción de carbohidratos, y que comparten una arquitectura básica común (Tabla 1). Tabla 1. Contenido de proteínas, lípidos y carbohidratos de membranas biológicas (expresado como porcentaje en peso). Membrana Proteína Lípido Carbohidratos Cociente proteína/lípido Eritrocito 49% 43% 8% 1.14 Célula hepática 54% 36% 10% 1.50 Membrana mitocondrial interna 78% 22% 0% 3.54 Según el modelo de Singer y Nicholson (1972), la membrana celular se compone de una capa bimolecular de lípidos, en la cual a determinados intervalos se incluyen unidades proteicas que forman un mosaico con la doble capa lipídica. En la membrana de la célula eucariota se localizan tres tipos de lípidos: fosfolípidos, glucolípidos y colesterol. a. Fosfolípidos. Se encuentran en mayor abundancia en las membranas. Las membranas contienen muchas clases de fosfolípidos distintos, incluyendo tanto a los glicero-fosfolípidos basados en glicerol como a los esfingolípidos basados en esfingosina. Los glicerofosfolípidos más comunes son fosfatidilcolina, fosfatidiletanolamina, fosfatidilserina y fosfatidilinositol. El esfingolípido más común es la esfingomielina. Las clases, orígenes y proporciones relativas de los fosfolípidos presentes en las membranas varían significativamente dependiendo del origen de éstas. Por ejemplo, la esfingomielina es uno de los principales fosfolípidos de las membranas plasmáticas animales, pero está ausente en las membranas plasmáticas de bacterias y plantas. La estructura de los fosfolípidos es la responsable de la actuación de las membranas como barreras entre dos compartimentos acuosos. Debido a que el interior de la bicapa fosfolipídica está ocupado por cadenas de ácidos grasos hidrofóbicos, la membrana es impermeable a moléculas hidrosolubles y a los iones. Los ácidos grasos de la mayor parte de los fosfolípidos naturales tienen uno o más enlaces dobles, que introducen flexión en las cadenas hidrocarbonadas y dificultan su empaquetamiento. Por tanto, las largas cadenas hidrocarbonadas de los ácidos grasos se mueven libremente en el interior de la membrana, por lo que la membrana es ligera y flexible. Además, los fosfolípidos como las proteínas son libres de difundir lateralmente dentro de la membrana -una propiedad fundamental para muchas de las funciones de la membrana. b. Glucolípidos. Se forman al añadir a los lípidos grupos carbohidrato. La mayoría de glucolípidos derivan de la esfingosina y, por tanto, se llaman glucoesfingolípidos. Los ejemplos más comunes son los cerebrósidos y los gangliósidos. Los cerebrósidos se denominan glucolípidos neutros ya que cada molécula tiene un azúcar no cargado como grupo principal -galactosa-, en el caso del galactocerebrósido. Por otro lado, un gangliósido, siempre tiene una cadena oligosacárida que contiene uno o más residuos de ácido siálico cargados negativamente, que le proporcionan a la molécula carga neta negativa. Los cerebrósidos y los gangliósidos predominan especialmente en las membranas cerebrales y en las células nerviosas. Los gangliósidos expuestos en la superficie de la membrana plasmática también funcionan como antígenos que son reconocidos por anticuerpos en
- 6. BIOLOGIA CELULAR 6 Seminario 1: Transporte a través de la Membrana 2020 – 20 las reacciones inmunes, incluyendo las reacciones responsables de las interacciones entre grupos sanguíneos. Por ejemplo, los grupos sanguíneos humanos ABO, implican a los glucoesfingolípidos que sirven como marcadores de la superficie celular de los eritrocitos. c. Esteroles. Las membranas de la mayoría de las células eucariotas contienen cantidades significativas de esteroles. El principal esterol de las membranas celulares animales es el colesterol. No se han encontrado esteroles en las membranas de las células procariotas. El colesterol contribuye a que la doble capa lipídica sea menos fluida. Esto se debe al rígido sistema cíclico esteroide de las moléculas de colesterol, que se interpone entre las mitades externas de las colas de los fosfolípidos. Las moléculas de colesterol también impiden que la viscosidad disminuya al descender la temperatura, dado que no permite el empaquetamiento denso (la cristalización) de las cadenas de ácidos grasos. Figura 1. Representación esquemática de los lípidos componentes de la bicapa lipídica). 2. Qué tipo de proteínas se encuentran en la membrana plasmática? Cuál es la función que cumplen las proteínas en las membranas celulares ? Dependiendo el tipo celular y el organelo en particular dentro de esa célula, una membrana puede contener cientos de proteínas diferentes. Cada proteína de la membrana posee una orientación definida en relación con el citoplasma para que las propiedades de una superficie de la membrana sean muy distintas respecto de las de otra superficie. Esta asimetría se conoce como “lateralidad” de la membrana. Por ejemplo, en la membrana plasmática las partes de las proteínas de membrana que interactúan con otras células o con ligandos extracelulares se proyectan hacia fuera, al espacio extracelular, en tanto que las partes de las proteínas de membrana que interactúan con moléculas del citoplasma se proyectan hacia el citosol. Las proteínas de membrana pueden agruparse en tres clases distintas que se distinguen por su estrecha relación con la bicapa lipídica. Estas son las siguientes: a. Proteínas integrales. Las proteínas integrales son proteínas transmembranosas, esto es, que cruzan toda la bicapa de lípidos y poseen, por tanto, 3 dominios: citosólico, trans-membrana y extracelular (o intraluminal en el caso de membranas de organelas). Algunas proteínas integrales tienen solo un segmento que abarca la membrana (unipaso), mientras que otras la cruzan varias veces (multipaso).
- 7. BIOLOGIA CELULAR 7 Seminario 1: Transporte a través de la Membrana 2020 – 20 Al igual que los fosfolípidos de la bicapa, las proteínas integrales de membrana también son anfipáticas y tienen porciones hidrófobas e hidrofílicas. Las porciones de una proteína integral de membrana que residen dentro de la bicapa lipídica tienden a tener un carácter hidrófobo. Los residuos de aminoácidos de estos dominios transmembranosos forman interacciones de van der Waals con las cadenas acilo grasas de la bicapa, lo cual sella a la proteína dentro de la “pared” de lípidos de la membrana. Como resultado, se conserva la barrera permeable y la proteína queda en contacto directo con las moléculas de lípido circundantes. b. Proteínas periféricas. Se localizan en su totalidad fuera de la bicapa de lípidos, ya sea en el lado citoplásmico o extracelular, aunque se relacionan con la superficie de la membrana mediante enlaces no covalentes (enlaces electrostáticos débiles). Por lo general, las proteínas periféricas pueden solubilizarse mediante la extracción con soluciones salinas en altas concentraciones que debilitan los enlaces electrostáticos que mantienen las proteínas periféricas en una membrana. Las proteínas periféricas mejor estudiadas se localizan en la superficie citosólica de la membrana plasmática, donde forman una red fibrilar que actúa como “esqueleto” de la membrana. Estas proteínas brindan soporte mecánico a la membrana y funcionan como un ancla para las proteínas integrales. c. Proteínas fijadas con lípidos (proteínas ancladas a la bicapa). Se localizan fuera de la bicapa de lípidos, ya sea en la superficie extracelular o la citosólica, pero que tienen enlaces covalentes con una molécula de lípidos que se sitúa dentro de la bicapa. Muchas proteínas presentes en la superficie externa de la membrana plasmática están unidas a esta mediante un pequeño oligosacárido complejo unido con una molécula de fosfatidilinositol que esta incrustada en la hoja externa de la bicapa lipídica. Las proteínas periféricas de membrana que contienen este tipo de enlace glucosilfosfatidilinositol se llaman proteínas fijadas con GPI. Otro grupo de proteínas presente en el lado citosólico de la membrana plasmática está fijado a la membrana mediante una o más cadenas largas de hidrocarburos embebidas en la hoja interna de la bicapa lipídica. Estos anclajes (restos palmitilo, miristilo, farnesilo y geranilgeranilo) definen las llamadas proteínas preniladas (p. ej. Ras, Rab, flotilina). La proteína ras está relacionada con la transformación de una célula normal en una maligna Figura 2. Proteínas de membrana. Las proteínas de membrana participan en las siguientes funciones: ♦ Transporte de solutos. Las proteínas funcionan transportando solutos a través de ellas. Entre éstas se incluyen las permeasas, que facilitan el movimiento de nutrientes como azúcares y aminoácidos
- 8. BIOLOGIA CELULAR 8 Seminario 1: Transporte a través de la Membrana 2020 – 20 a través de las membranas, las proteínas canal que proporcionan vías de paso hidrofílicas a través de membranas hidrofóbicas y ATPasas transportadoras, que utilizan la energía del ATP para bombear iones a través de las membranas. ♦ Enzimas. Las enzimas de membrana catalizan reacciones químicas específicas que ocurren en la superficie externa de la célula o apenas dentro de la célula. Ej. Las enzimas ubicadas sobre la superficie externa de las células que recubren el intestino delgado son responsables de la digestión de carbohidratos y péptidos. Las enzimas unidas a la superficie intracelular de muchas membranas celulares desempeñan un papel importante en la transferencia de señales desde el ambiente externo extracelular hacia el citoplasma. ♦ Receptores. Están implicados en el reconocimiento y mediación de los efectos de señales químicas específicas que se unen a la superficie de la célula. Las hormonas, neuro-transmisores y sustancias que promueven el crecimiento son ejemplos de señales químicas que interactúan con receptores proteicos específicos situados en la membrana plasmática de sus células diana. En la mayoría de los casos, la unión de una hormona o de otra molécula señal al receptor apropiado de la superficie de la membrana desencadena un tipo de respuesta intracelular, para obtener el efecto deseado. ♦ Adhesión y comunicación entre células adyacentes. La mayoría de las células de los organismos multicelulares están en contacto con otras células, a través de conexiones citoplasmáticas directas, que permiten, al menos, el intercambio de algún componente celular. Esta comunicación intercelular viene proporcionada por las uniones gap en las células animales y por los plasmodesmos de las células vegetales. ♦ Estructural. Las proteínas estructurales tienen tres papeles principales. El primero de ellos es el de conectar la membrana con el citoesqueleto para mantener la forma de la célula. El segundo papel es el de crear uniones celulares que mantengan unidos a los tejidos, como las uniones estrechas y las uniones en hendidura. Finalmente, el tercer papel es el de fijar las células a la matriz extracelular mediante la conexión de fibras del citoesqueleto con la fibras extracelulares de colágeno y otras proteínas. . Figura 3. Funciones de la proteínas de membrana.
- 9. BIOLOGIA CELULAR 9 Seminario 1: Transporte a través de la Membrana 2020 – 20 3. Cómo se definen gradiente químico y gradiente eléctrico ? Qué tipos de transporte ocurre en las membranas celulares ?. Cómo se diferencian cada uno de ellos ? Gradiente químico, es cuando una sustancia está presente en diferentes concentraciones a ambos lados de la membrana. El término gradiente de concentración puede considerarse como sinónimo de diferencia de concentración Estrictamente hablando, el término gradiente de concentración se usa en relación con cualquier diferencia en la concentración entre una localización y otra, no solo diferencias entre un lado y otro de la membrana. Cuando las moléculas se mueven de mayor a menor concentración, podemos decir que se están moviendo a favor de su gradiente de concentración; el movimiento en la dirección contraria es en contra del gradiente de concentración. Gradiente eléctrico, es la diferencia de voltaje (potencial eléctrico) entre ambos lados de la membrana. El gradiente eléctrico afecta la difusión de todos los solutos con carga eléctrica. Figura 4. Tipos de transporte a través de la membrana. El transporte de sustancias a través de las membranas celulares se realiza mediante varios tipos de mecanismos. Las sustancias pueden ser transportadas a favor de un gradiente electroquímico (descendente) o en contra de un gradiente electroquímico (ascendente). El transporte descendente se produce por difusión simple o facilitada y no requiere ningún aporte de energía metabólica. El transporte ascendente se produce por transporte activo, que puede ser primario o secundario. El transporte activo primario y secundario se distinguen entre sí por la fuente de energía. El primero requiere un aporte directo de energía metabólica, mientras que el segundo utiliza un aporte indirecto de energía metabólica. Otras diferencias entre los mecanismos de transporte se basan en si el proceso incluye una proteína transportadora. La difusión simple es la única forma de transporte que no está mediada por una proteína transportadora. En la difusión facilitada, en el transporte activo primario y el secundario intervienen proteínas integrales de membrana y se denominan transporte mediado por transportador. La difusión facilitada tiene las mismas propiedades que la difusión simple, las moléculas transportadas se mueven a favor de su gradiente de concentración, el proceso no requiere aporte de energía y el movimiento neto se detiene en el equilibrio, cuando la concentración en el interior de la célula se iguala con la concentración en el exterior.
- 10. BIOLOGIA CELULAR 10 Seminario 1: Transporte a través de la Membrana 2020 – 20 La difusión facilitada difiere de la difusión simple en la siguiente característica importante: aunque la velocidad de la difusión simple a través de un canal abierto aumenta de manera proporcional a la concentración de la sustancia que difunde, en la difusión facilitada la velocidad de difusión se acerca a un máximo, denominado Vmáx, a medida que aumenta la concentración de la sustancia que difunde. Tabla 2. Características de los procesos de transporte. 4. Cuál es la composición química de los líquidos intracelular y extracelular? Las composiciones del Líquido Intracelular (LIC) y del Líquido extracelular (LEC) son notablemente diferentes, como se muestra en la tabla 2. El principal catión del LEC es el sodio (Na+ ) y los aniones de equilibrio son cloro (Cl− ) y bicarbonato (HCO3− ). Los principales cationes del LIC son potasio (K+ ) y magnesio (Mg2+ ), y los aniones de equilibrio son proteínas y fosfatos orgánicos. Otras diferencias significativas de composición afectan al Ca2+ y al pH. Habitualmente, el LIC tiene una concentración muy baja de Ca2+ ionizado (≈10−7 mol/l), mientras que la concentración de Ca2+ en el LEC es mayor, de aproximadamente 4 órdenes de magnitud. El LIC es más ácido (tiene un pH más bajo) que el LEC. Por tanto, las sustancias que se encuentran en concentraciones altas en el LEC tienen concentraciones bajas en el LIC y viceversa. Sorprendentemente, dadas las diferencias de concentración de solutos individuales, la concentración total de solutos (osmolaridad) es igual en el LIC y el LEC. Esta equivalencia se consigue porque el agua fluye libremente a través de las membranas celulares. Cualquier diferencia transitoria de la osmolaridad que haya entre el LIC y el LEC se disipa rápidamente por el movimiento del agua hacia el interior o el exterior de las células para restablecer la equivalencia. Las diferencias en la concentración de solutos a través de las membranas celulares se crean y se mantienen mediante mecanismos de transporte con gasto de energía en las membranas celulares. El más conocido de estos mecanismos de transporte es la Na+ /K+ ATPasa (bomba de Na+ /K+ ), que transporta Na+ del LIC al LEC y, simultáneamente, transporta K+ del LEC al LIC. Tanto el Na+ como el K+ se transportan contra sus gradientes electroquímicos respectivos; por lo que necesitan de adenosina trifosfato (ATP). La Na+ /K+ ATPasa se encarga de crear los grandes gradientes de concentración de Na+ y K+ que se producen a través de las membranas celulares (es decir, la baja concentración intracelular de Na+ y la alta concentración intracelular de K+ ).
- 11. BIOLOGIA CELULAR 11 Seminario 1: Transporte a través de la Membrana 2020 – 20 Tabla 3. Comparación de la concentración media de varias moléculas en una célula de mamífero De forma parecida, la concentración intracelular de Ca2+ se mantiene a un nivel mucho más bajo que la concentración extracelular de Ca2+ . Esta diferencia de concentración se establece, en parte, por una Ca2+ ATPasa de la membrana celular que bombea Ca2+ en contra de su gradiente electroquímico, con gasto de ATP. Además de los transportadores que utilizan directamente ATP, otros transportadores establecen diferencias de concentración a través de la membrana celular utilizando el gradiente de concentración transmembrana de Na+ . Estos transportadores crean gradientes de concentración para glucosa, aminoácidos, Ca2+ e H+ sin la utilización directa de ATP. Directa o indirectamente, las diferencias de composición entre el LIC y LEC se encuentran en cada función fisiológica importante, como ilustran los siguientes ejemplos: a. El potencial de membrana en reposo del nervio y el músculo depende fundamentalmente de la diferencia en la concentración de K+ a través de la membrana celular; b. El aumento del potencial de acción de estas células excitables depende de las diferencias en la concentración de Na+ a través de la membrana celular; c. El acoplamiento excitación-contracción en las células musculares depende de las diferencias en la concentración de Ca2+ a través de la membrana celular y la membrana del retículo sarcoplásmico, d. La absorción de nutrientes esenciales depende del gradiente de concentración transmembrana de Na+ (p. ej., absorción de glucosa en el intestino delgado o reabsorción de glucosa en el túbulo renal proximal). 5. Qué es la difusión simple ?. Qué factores pueden afectar la difusión ? La difusión (diffundere, derramarse un líquido) puede definirse como el movimiento pasivo de las moléculas no cargadas desde un área de mayor concentración hacia un área de menor concentración debido al movimiento molecular al azar. Como resultado, habrá un movimiento neto desde la región de concentración más alta hacia la de concentración más baja hasta que la diferencia de concentración ya no exista. Este movimiento neto se llama difusión neta. La difusión neta es un proceso físico que ocurre siempre que hay una diferencia de concentración a través de la membrana, y la membrana es permeable a la sustancia que esté difundiendo. Por ejemplo, la concentración de oxigeno (O2) es relativamente alta en el líquido extracelular porque la sangre transporta el oxígeno desde los pulmones hacia los tejidos corporales. Puesto que el oxígeno se combina con hidrogeno para formar agua en la respiración celular aeróbica, la concentración de oxigeno dentro de las células es más baja que en el líquido extracelular. Los eritrocitos del sistema circulatorio tomen oxígeno en los pulmones y lo liberen luego en los tejidos. En los capilares de los tejidos corporales, donde la concentración de oxígeno es baja, éste es liberado desde la hemoglobina y difunde
- 12. BIOLOGIA CELULAR 12 Seminario 1: Transporte a través de la Membrana 2020 – 20 pasivamente desde el citoplasma del eritrocito hacia el plasma sanguíneo y desde allí a las células que limitan los capilares. En los capilares pulmonares se verifica el proceso contrario: el oxígeno difunde desde el aire inhalado en los pulmones, donde su concentración es mayor, al citoplasma de los eritrocitos, donde la concentración es menor. El dióxido de carbono también atraviesa las membranas por difusión simple; sin embargo, la mayoría del CO2 viaja en forma de ion bicarbonato (HCO3- ). No es de extrañar que el dióxido de carbono y el oxígeno atraviesen la membrana en sentido opuesto, Los factores que afectan a la difusión simple son: a. Tamaño del soluto. En términos generales, las bicapas lipídicas son más permeables a las moléculas pequeñas que a las grandes. Las moléculas pequeñas más relevantes para la función celular son el agua, el oxígeno y el dióxido de carbono. Las membranas son bastante permeables a estas moléculas; no se requieren mecanismos de transporte específicos para introducirlas o sacarlas de las células. Aunque el agua no es liposoluble, las moléculas de agua pueden difundirse a través de la membrana plasmática hasta un grado limitado debido a su pequeño tamaño (difunden a través de una bicapa, 10.000 veces más despacio de lo que se mueven en ausencia de una membrana) y carencia de carga neta. La regla del tamaño sirve para moléculas de hasta aproximadamente unos cien daltons. El etanol (CH3CH2OH; 46 daltons) y el glicerol (C3H8O3; 92 daltons) son capaces de difundir a través de membranas con una tasa de transporte razonable, pero no así la glucosa (C6H12O6; 180 daltons). b. Polaridad del soluto. En general las bicapas lipídicas son relativamente permeables a moléculas no polares y menos permeables a las moléculas polares. Esto es debido a que las moléculas no polares se disuelven más fácilmente en la fase hidrófoba de la bicapa lipídica y así pueden atravesar la membrana mucho más rápidamente que las moléculas polares de tamaño similar. Si a un compuesto polar como la úrea, que presenta una permeabilidad relativamente baja, se le añaden dos grupos metilo para formar dimetilurea, se puede disminuir su polaridad, por lo tanto, aumentar su permeabilidad. La relativa impermeabilidad a las sustancias polares en general y a los iones en particular, es debida a su fuerte asociación con las moléculas de agua, que forman un escudo de hidratación. Para mover tales solutos en la membrana, se requiere que sean desnudados del agua, eliminando los enlaces entre los iones y las moléculas de ésta. La asociación de los iones con las moléculas de agua para formar los escudos de hidratación es un fenómeno que restringe dramáticamente el transporte de iones en las membranas. Las moléculas polares de mayor tamaño, como la glucosa, no pueden pasar a través de la doble capa de moléculas de fosfolípidos y, así, requieren proteínas transportadoras especiales en la membrana para su transporte. La porción fosfolípido de la membrana es similarmente impermeable a iones inorgánicos cargados, como Na+ y K+ . De cualquier modo, pequeños canales iónicos permiten el paso de estos iones. La impermeabilidad de las membranas a iones es muy importante para la actividad celular, puesto que todas las células deben mantener un potencial electroquímico de membrana para su correcto funcionamiento. En la mayoría de los casos se consigue con un gradiente de sodio (células animales) o de protones (casi todas las demás células). c. La difusión está relacionada con la temperatura. A temperatura elevada, las moléculas se mueven más rápidamente. Puesto que la difusión es resultado del movimiento molecular, la velocidad de difusión aumenta a medida que lo hace la temperatura. Generalmente, los cambios de temperatura no afectan significativamente las velocidades de difusión en el hombre, ya que éste mantiene una temperatura corporal relativamente constante.
- 13. BIOLOGIA CELULAR 13 Seminario 1: Transporte a través de la Membrana 2020 – 20 d. La velocidad de difusión depende de la capacidad de la molécula que difunde de disolverse en la bicapa lipídica de la membrana. Las moléculas difieren respecto a su capacidad de disolverse en lípidos. Las sustancias hidrófobas (no polares) son las más solubles en lípidos, mientras que las hidrófilas (polares o cargadas) son las menos solubles. Dado que las moléculas deben moverse a través de una bicapa lipídica en la difusión simple, cabe esperar que las moléculas liposolubles sean capaces de atravesar la bicapa más fácilmente. En igualdad de condiciones, cuanto más liposoluble sea una sustancia, más permeabilidad tendrá la membrana a esa sustancia. La liposolubilidad también está influenciada por la composición lipídica de la membrana, que varía ligeramente de una membrana a otra. Las membranas con alto contenido de colesterol son menos permeables al agua que aquellas con bajo contenido de colesterol, presumiblemente porque las moléculas de colesterol ocupan los espacios entre las colas de ácidos grasos de la bicapa lipídica y, así, excluyen al agua. Debido a que la mayoría de sustancias en el organismo son hidrófilas y, por tanto, no atraviesan la bicapa lipídica fácilmente, la lista de sustancias conocidas que puedan ser transportadas por difusión simple es muy limitada, e incluye ácidos grasos, hormonas esteroideas, hormonas tiroideas, oxígeno, dióxido de carbono y las vitaminas liposolubles (A, D, E y K). e. La velocidad de difusión es inversamente proporcional al espesor de la membrana. Mientras mayor espesor tenga la membrana más lenta será la velocidad a la cual ocurre la difusión. En la mayoría de las membranas biológicas, el espesor es esencialmente constante; pero, el espesor de los tejidos varía considerablemente. Los tejidos especia-lizados en el transporte, como las paredes de los capilares o del epitelio pulmonar, tienden a tener las pare des relativamente finas; esta delgadez aumenta la permeabilidad y mejora la velocidad de transporte a través de estos tejidos. Figura 5. Permeabilidad de las bicapas fosfolipídicas.
- 14. BIOLOGIA CELULAR 14 Seminario 1: Transporte a través de la Membrana 2020 – 20 6. Qué es la difusión facilitada ?. Qué características tienen los transportadores ? La difusión facilitada también se denomina difusión mediada por un transportador porque una sustancia que se transporta de esta manera difunde a través de la membrana utilizando una proteína transportadora específica para contribuir al transporte. Es decir, el transportador facilita la difusión de la sustancia hasta el otro lado. Los transportadores son proteínas complejas, grandes y con múltiples subunidades. El cambio de conformación que se exige a una proteína transportadora hace a esta forma de transporte mucho más lenta que el movimiento a través de los canales proteicos. Todas las formas de transporte mediado por transportador comparten tres características: saturación, estereoespecificidad y competición. El transporte mediante proteínas transportadoras se caracteriza por lo siguiente: ♦ Estereoespecificidad. Los sitios de unión para el soluto en las proteínas transportadoras son estereoespecíficos. Por ejemplo, el transportador de glucosa en el túbulo proximal renal reconoce y transporta el isómero natural (la D-glucosa) pero no reconoce ni transporta el isómero no natural (la L-glucosa). En cambio, la difusión simple no distingue entre los dos isómeros de glucosa porque no interviene ninguna proteína transportadora. ♦ Saturación. La saturabilidad se basa en el concepto de que las proteínas portadoras tienen un número limitado de sitios de unión para el soluto. En concentraciones bajas de solutos, existen muchos sitios de unión y la velocidad del transporte aumenta considerablemente a medida que aumenta la concentración. Sin embargo, en concentraciones altas de solutos, apenas hay sitios de unión y la velocidad de transporte se estabiliza. Por último, cuando todos los sitios de unión están ocupados, se alcanza la saturación en un punto llamado transporte máximo o Tm. La cinética del transporte mediado por portador es similar a la cinética enzimática de Michaelis-Menten; en ambas intervienen proteínas con un número limitado de sitios de unión. ♦ Competición. Aunque los sitios de unión de los solutos transportados son bastante específicos, pueden reconocer, unir e incluso transportar solutos químicamente relacionados. Por ejemplo, el transportador de glucosa es específico de la D-glucosa, pero también reconoce y transporta un azúcar muy relacionado (la D-galactosa). Por tanto, la presencia de D-galactosa inhibe el transporte de D-glucosa al ocupar algunos de los sitios de unión, haciendo que no estén disponibles para la glucosa. ♦ Reversibilidad según el gradiente. El transporte es bidireccional, de modo que la dirección se determina dependiendo del gradiente de concentración a ambos lados de la membrana. 7. Que es un canal iónico? Qué tipos de canales existen? Los canales iónicos son canales proteicos constituidos por subunidades proteicas que atraviesan la membrana y crean un grupo de cilindros que rodean a un poro angosto, lleno de agua. Cuando los canales están abiertos, decenas de millones de iones por segundo pueden pasan rápida y libremente a través de ellos, moviéndose a favor del gradiente electroquímico. Los canales iónicos son selectivos y permiten el paso de iones con características específicas. Esta selectividad está determinada por el diámetro de su poro central y por la carga eléctrica de los aminoácidos que la recubren. Por ejemplo, los canales revestidos de cargas negativas habitualmente permiten el paso de cationes, pero excluyen los aniones; los canales revestidos de cargas positivas permiten el paso de aniones, pero excluyen los cationes. Los canales también discriminan según el tamaño. Por ejemplo, un canal selectivo para los cationes revestido de cargas negativas permitirá el paso de Na+ , pero excluirá el K+ ; otro canal selectivo para los cationes (p. ej., un receptor nicotínico en la placa motora terminal) podría tener menor selectividad y
- 15. BIOLOGIA CELULAR 15 Seminario 1: Transporte a través de la Membrana 2020 – 20 permitir el paso de pequeños cationes diferentes. Los canales pueden clasificarse en los que tienen sus compuertas generalmente abiertas o generalmente cerradas. Los canales abiertos pasan la mayor parte del tiempo con su compuerta abierta, lo que permite que sus iones se desplacen a través de la membrana en ambos sentidos sin regulación. Los canales tipo compuerta pasan la mayor parte del tiempo cerrados, lo que les permite regular el movimiento de los iones a través de ellos. Cuando se abre un canal tipo compuerta, los iones se desplazan a través del canal de la misma manera que lo hacen a través de los canales abiertos, Cuando un canal tipo compuerta está cerrado, como lo está la mayor parte del tiempo, no permite el movimiento de iones entre los líquidos intracelular y extracelular. Los tipos de canales son: ♦ Los canales dependientes del ligando. Las compuertas de algunos canales proteicos se abren por la unión de una sustancia química (un ligando: hormonas, neurotransmisores y segundos mensajeros) a la proteína; esto produce un cambio conformacional o un cambio de los enlaces químicos de la molécula de la proteína que abre o cierra la compuerta. El ligando a menudo es externo (p. ej., un neurotransmisor u hormonas). Sin embargo, también puede ser interno; Ca2+ intracelular, monofosfato de adenosina cíclico (cAMP). Uno de los casos más importantes de activación química es el efecto de la acetilcolina sobre el denominado canal de la acetilcolina. La acetilcolina abre la compuerta de este canal, dando lugar a la apertura de un poro de carga negativa que permite que lo atraviesen moléculas sin carga o iones positivos menores de este diámetro. Esta compuerta es muy importante para la transmisión de las señales nerviosas desde una célula nerviosa a otra y desde las células nerviosas a las células musculares para producir la contracción muscular. ♦ Los canales dependientes del voltaje. La conformación molecular de la compuerta o de sus enlaces químicos responde al potencial eléctrico que se establece a través de la membrana celular. Por ejemplo, la compuerta de activación en el canal de Na+ , cuando hay una carga negativa intensa en el interior de la membrana celular, hace que las compuertas de sodio del exterior permanezcan firmemente cerradas; por el contrario, cuando el interior de la membrana pierde su carga negativa estas compuertas se abrirían súbitamente y permitirían que cantidades muy grandes de sodio entraran a través de los poros de sodio. Este es el mecanismo básico para generar los potenciales de acción nerviosos que son responsables de las señales nerviosas. Es de destacar que otra compuerta en el canal de Na+ , una compuerta de inactivación, se cierra por despolarización. Figura 6. Mecanismos de pasaje de los iones a través de los canales iónicos. ♦ Los conductos mecanosensitivos. Cuyo estado de conformación depende de fuerzas mecánicas (p.ej. estiramiento) que se aplican a la membrana. Por ejemplo, los miembros de una familia de conductos catiónicos se abren por el movimiento de estereocilios en las células vellosas del oído interno como respuesta al sonido o movimiento de la cabeza.
- 16. BIOLOGIA CELULAR 16 Seminario 1: Transporte a través de la Membrana 2020 – 20 8. Cómo se produce el transporte de glucosa por difusión facilitada?. La glucosa es la principal fuente corporal de energía directa y la mayor parte de los mamíferos contienen una proteína de membrana que facilita la difusión de la glucosa del torrente sanguíneo a la célula. La proteína transportadora tiene un sitio que se une de manera específica a la glucosa, y esa unión causa un cambio conformacional en el transportador, de modo que se forma una vía a través de la membrana. Como resultado, se permite que la glucosa se difunda a favor de su gradiente de concentración hacia la célula. Los transportadores para la difusión facilitada de glucosa se designan con las letras GLUT, seguidas por un número para la isoforma. El transportador para la glucosa en eritrocitos, se designa GLUT1; en el hígado es el GLUT2, en músculos esqueléticos es el GLUT4. La concentración de glucosa suele ser mayor en el medio extracelular (sangre en el caso de los eritrocitos) que en la célula, el GLUT1 cataliza la entrada neta de glucosa desde el medio extracelular hacia el interior de la célula. En estas condiciones, la V máx. se alcanza a altas concentraciones externas de glucosa. Se supone que GLUT1 transporta la glucosa por un mecanismo de alternancia de conformaciones. Los dos estados conformacionales se denominan T1, con el lugar de unión de glucosa abierto hacia el exterior de la célula y T2, que se abre hacia el interior celular. El proceso se inicia cuando una molécula de D-glucosa colisiona y se une a una molécula de GLUT1, que está en su conformación T1. Con la glucosa unida, GLUT1 cambia ahora a su conformación T2. El cambio conformacional permite la liberación de la molécula de glucosa hacia el interior de la célula, después de lo cual, la molécula de GLUT1 vuelve a su conformación inicial, es decir con el lugar de unión abierto de nuevo hacia el exterior. Figura 7. Modelo de conformación alternante para la difusión facilitada de glucosa, en GLUT1 El proceso es reversible, puesto que las permeasas funcionan en ambos sentidos. Las moléculas de soluto pueden entrar o salir, dependiendo de su concentración relativa a ambos lados de la membrana. Si la concentración es mayor en el exterior, el flujo neto será de entrada; si por el contrario la concentración es mayor en el interior, el flujo será de salida. El GLUT1 es el 2% de las proteínas en la membrana plasmática de los eritrocitos. Después que la glucosa es transportada al interior del eritrocito, es rápidamente fosforilada y forma glucosa 6-fosfato, que no puede abandonar la célula (GLUT1, como la mayoría de los transportadores de azúcares, no reconoce a las formas fosforiladas). Esto permite que la concentración intracelular de glucosa no se incremente a medida que la glucosa es incorporada por la célula.
- 17. BIOLOGIA CELULAR 17 Seminario 1: Transporte a través de la Membrana 2020 – 20 9. Qué es ósmosis ?. Cómo responde un eritrocito frente a distintos tipos de soluciones ? La ósmosis es la difusión neta de agua (el solvente) a través de la membrana. Para que ocurra osmosis, la membrana debe ser selectivamente permeable; es decir, debe ser más permeable a moléculas de agua que a por lo menos una especie de soluto. Así, hay dos requerimientos para la osmosis: a) debe haber una diferencia en la concentración de un soluto en los dos lados de una membrana selectivamente permeable, y b) la membrana debe ser relativamente impermeable al soluto. Los solutos que no pueden pasar de manera libre a través de la membrana pueden promover el movimiento osmótico de agua, y se dice que son osmóticamente activos. La difusión de agua ocurre cuando el agua está más concentrada en un lado de la membrana que en el otro lado; es decir, cuando una solución está más diluida que la otra. Es decir, hay un movimiento neto de moléculas de agua desde el lado de concentración más alta de agua hacia el lado de concentración más baja. El término tonicidad se usa para describir el efecto de una solución sobre el movimiento osmótico de agua; por ejemplo, si una solución glucosada o salina isoosmótica está separada del plasma por una membrana que es permeable al agua, pero no a la glucosa o al NaCl, no ocurrirá osmosis. En este caso, se dice que la solución es isotónica (del griego isos, “igual”; tonos, “tensión”) respecto al plasma. Dado que la dextrosa al 5% (5 g de glucosa por 100 ml, que es 0.3 m) y la solución salina normal (0.9 g de NaCl por 100 ml, que es 0.15 m) tienen la misma osmolalidad que el plasma, se dice que son isoosmóticas respecto al plasma. Debemos hacer notar que una solución puede ser isoosmótica pero no isotónica; tal es el caso siempre que el soluto en la solución isoosmótica puede penetrar libremente la membrana; por ejemplo, una solución de úrea 0.3 m es isoosmótica pero no isotónica porque la membrana celular es permeable a la urea. Los eritrocitos en una solución de úrea 0.3 m estallarán. Un medio o solución isotónica es aquel en donde la concentración de soluto en el medio exterior en relación al medio interior de la célula es igual tanto en el exterior como en el interior de una célula. Al tener la misma concentración tendrán aproximadamente la misma presión osmótica por lo que en ese medio, la célula no sufrirá deformación física alguna. Los eritrocitos colocados en una solución isotónica no ganaran agua ni la perderán. Figura 7. Efectos de las diferencias en la concentración de solutos en la membrana plasmática Las soluciones que tienen una concentración total más baja de solutos que la del plasma, y que por consiguiente tienen una presión osmótica más baja, son hipoosmóticas respecto al plasma. Los eritrocitos colocados en soluciones hipotónicas ganan agua y pueden estallar, proceso llamado hemólisis. Cuando los eritrocitos se colocan en una solución hipertónica (como agua de mar), que contiene solutos
- 18. BIOLOGIA CELULAR 18 Seminario 1: Transporte a través de la Membrana 2020 – 20 osmóticamente activos a una osmolalidad y presión osmótica más altas que las del plasma, disminuyen de tamaño debido a la ósmosis de agua hacia afuera de las células. Este proceso se llama crenación (del latin crena, “muesca”) porque la superficie de la célula adopta un aspecto festoneado. La ósmosis es un factor importante en una multitud de funciones corporales. Por ejemplo, el aparato digestivo secreta varios litros de líquido al día, que se reabsorben por ósmosis a través de las células que recubren el intestino. Si este líquido no se absorbiera, como ocurre en casos de diarrea extrema, ocurrirá una deshidratación rápida. 10. Qué es transporte activo ?. Diferencie los distintos tipos de transportadores. El transporte activo es el movimiento de moléculas e iones contra sus gradientes de concentración, desde concentraciones más bajas hacia más altas. Es decir, el soluto se mueve de una zona de baja concentración (o potencial electroquímico bajo) a otra de alta concentración (o potencial electroquímico alto). Puesto que el movimiento ascendente de un soluto es un trabajo, debe aportarse energía metabólica en forma de ATP. Dado que el transporte activo comprende el transporte de iones y moléculas cuesta arriba, contra su gradiente de concentración, y usa energía metabólica, los transportadores de transporte activo primario se denominan bombas. Tabla 3. Principales tipos de ATPasa de transporte (bombas) El transporte activo primario ocurre cuando la hidrólisis de ATP es directamente responsable de la función de los transportadores, que son proteínas que abarcan el grosor de la membrana. Los siguientes son ejemplos de transporte activo primario en los sistemas fisiológicos: la Na+ /K+ ATPasa presente en todas las membranas celulares, la Ca2+ ATPasa presente en los retículos sarcoplásmico y endoplásmico y la H+ /K+ ATPasa de las células parietales del estómago (de la cual depende la acidez del jugo gástrico del estómago),
- 19. BIOLOGIA CELULAR 19 Seminario 1: Transporte a través de la Membrana 2020 – 20 Las estructuras de las bombas iónicas de clase F y clase V son similares entre sí, pero no están emparentadas con las bombas de clase P y son más complicadas que ellas. Las bombas de clases F y V contienen varias subunidades citosólicas y transmembrana diferentes. Todas las bombas V y F conocidas transportan sólo protones, en un proceso que no involucra una fosfoproteína intermediaria. Las bombas de clase V suelen cumplir funciones para mantener el pH bajo de las vacuolas de las plantas, de los lisosomas y de otras vesículas acídicas en las células animales al bombear protones desde la cara citosólica a la exoplasmática de la membrana en contra de un gradiente electroquímico de protones. Las bombas de clase F se encuentran en las membranas plasmáticas bacterianas, en la mitocondria y en los cloroplastos. A diferencia de las bombas de clase V, las de clase F suelen funcionar para impulsar la síntesis de ATP a partir de ADP y Pi, a través del movimiento de protones desde la cara exoplasmática hacia la citosólica de la membrana a favor del gradiente electroquímico de protones. Debido a su importancia en la síntesis de ATP en los cloroplastos y en la mitocondria, las bombas de protones de clase F, son llamadas ATP sintasas. 11. Qué características tienen las bombas de tipo P?. Cómo funciona la bomba de sodio/potasio ?. Porqué se dice electrogénica ? ATPasas de tipo P. Las ATPasas de tipo P (P de «phosphorylation ») se fosforilan reversiblemente por ATP, en residuos de aspartato, durante el transporte. La mayor parte de las bombas de tipo P se localizan en la membrana celular, donde son las responsables del mantenimiento del gradiente iónico a ambos lados de la membrana plasmática. Todas las bombas de iones de clase P poseen dos subunidades α catalíticas que contienen un sitio fijador de ATP. La mayoría también tiene dos subunidades β más pequeñas que cumplen funciones reguladoras. Durante el proceso de transporte, al menos una de las subunidades α es fosforilada y se piensa que los iones transportados se mueven a través de la subunidad fosforilada. Esta clase incluye la Na+ /K+ ATPasa de la membrana plasmática, las Ca2+ ATPasas y la H+ /K+ ATPasa. La Na+ /K+ ATPasa se encuentra en las membranas de todas las células. Es un complejo formado por dos proteínas globulares distintas: una de mayor tamaño denominada subunidadα α, que tiene un peso molecular de aproximadamente 100.000, y una más pequeña denominada subunidad β, que tiene un peso molecular de aproximadamente 55.000. La proteína de mayor tamaño tiene tres características específicas que son importantes para el funcionamiento de la bomba: a. Tiene tres puntos receptores para la unión de iones sodio en la porción de la proteína que protruye hacia el interior de la célula. b. Tiene dos puntos receptores para iones potasio en el exterior. c. La parte interior de esta proteína cerca de puntos de unión al Na+ tiene actividad ATPasa. La Na+ /K+ ATPasa bombea Na+ del LIC al LEC y K+ del LEC al LIC. Cada ion se mueve contra su gradiente electroquímico correspondiente. Generalmente, por cada 3 iones Na+ bombeados fuera de la célula, se bombean 2 iones K+ hacia dentro. Esta estequiometría de 3 iones Na+ por 2 iones K+ significa que, por cada ciclo de la Na+ /K+ ATPasa, se bombea más carga positiva fuera de la célula que hacia dentro. Por tanto, el proceso se denomina electrogénico porque crea una separación de cargas y una diferencia de potencial. La Na+ /K+ ATPasa se encarga de mantener los gradientes de concentración de Na+ y K+ a través de las membranas celulares, manteniendo baja la concentración intracelular de Na+ y alta la concentración intracelular de K+ . La función de transporte de iones (es decir, bombear Na+ fuera de la célula y K+ hacia dentro) se basa en el ciclo entre los estados E1 y E2 y la energía se obtiene de la hidrólisis del ATP. El ciclo de transporte empieza con la proteína transportadora en el estado E1, unida al ATP. En el estado E1, los sitios de unión de los iones están de cara al líquido intracelular y la proteína tiene una alta afinidad
- 20. BIOLOGIA CELULAR 20 Seminario 1: Transporte a través de la Membrana 2020 – 20 por el Na+ ; se unen 3 iones Na+ , el ATP es hidrolizado y el fosfato terminal del ATP es transferido al transportador produciendo un estado E1∼P, de alta energía. Ahora tiene lugar un gran cambio conformacional y la proteóna pasa de E1∼P a E2∼P. En el estado E2, los sitios de unión de los iones están de cara al líquido extracelular, la afinidad por el Na+ es baja y la afinidad por el K+ , alta. Los 3 iones Na+ se liberan de la proteína transportadora al líquido extracelular, se unen 2 iones K+ y se libera fosfato inorgánico de E2. La proteína ahora se une al ATP intracelular y hay otro gran cambio conformacional que devuelve la proteína al estado E1; los 2 iones K+ se liberan al líquido intracelular y la enzima está preparada para otro ciclo. Figura 8. Modelo de funcionamiento de la Na+ /K+ ATPasa. Una de las funciones más importantes de la bomba Na+ /K+ es controlar el volumen de todas las células. Sin la función de esta bomba la mayor parte de las células del cuerpo se hincharía hasta explotar. El mecanismo para controlar el volumen es el siguiente: en el interior de la célula hay grandes cantidades de proteínas y de otras moléculas orgánicas que no pueden escapar de la célula. La mayor parte de ellas tiene carga negativa y, por tanto, atrae grandes cantidades de potasio, sodio y también de otros iones positivos. Todas estas moléculas e iones producen ósmosis de agua hacia el interior de la célula. Salvo que este proceso se detenga, la célula se hinchará indefinidamente hasta que explote. El mecanismo normal para impedirlo es la bomba Na+ /K+ Los glucósidos cardíacos (o cardiotónicos; p. ej., ouabaína y digital) son una clase de fármacos que inhiben la Na+ /K+ ATPasa. El tratamiento con esta clase de fármacos genera ciertos cambios predecibles en la concentración iónica intracelular: la concentración intracelular de Na+ aumentará y la de K+ disminuirá. Los glucósidos cardíacos inhiben la Na+ /K+ ATPasa por unión a la forma E2∼P cerca del sitio de unión del K+ en el lado extracelular, evitando, por tanto, la conversión de E2∼P en E1. Al alterar el
- 21. BIOLOGIA CELULAR 21 Seminario 1: Transporte a través de la Membrana 2020 – 20 ciclo fosforilación-desfosforilación, estos fármacos alteran todo el ciclo enzimático y sus funciones de transporte. 12. Cómo funciona las otras bombas de tipo P: bomba de calcio e hidrógeno/potasio ? Las bombas de Ca2+ están ubicadas en la membrana plasmática (Ca2+ ATPasa de membrana plasmática; PMCA, del inglés plasma-membrane Ca2+ ATPase), de todas las células, y en la membrana del retículo endoplásmico (Ca2+ ATPasa del retículo sarcoplásmico y endoplásmico – SERCA) de células de musculo estriado y otras. Los iones calcio normalmente se mantienen a una concentración muy baja en el citosol intracelular de prácticamente todas las células del cuerpo. Esto se consigue principalmente mediante dos bombas de calcio que funcionan mediante transporte activo primario. Una está en la membrana celular que extrae un ion Ca2+ por cada ATP hidrolizado. La otra bombea 2 iones Ca2+ (por cada ATP hidrolizado) del líquido intracelular al interior del retículo sarcoplásmico o endoplásmico, es decir, un secuestro de Ca2+ . Debido al gradiente de concentración así creado, cuando canales de iones para Ca2+ se abren en la membrana plasmática o el retículo endoplasmático, el Ca2+ se difundirá con rapidez a favor de su gradiente de concentración hacia el citoplasma. Este aumento repentino del Ca2+ citoplasmático sirve como una señal para diversos procesos, entre ellos la liberación de neurotransmisores desde terminales de axón y contracción muscular. Figura 9. Modelo del mecanismo de acción de la funcionamiento de la Ca2+ ATPasa. En la membrana del retículo sarcoplásmico de las células del músculo esquelético La bomba de Ca2+ presenta un sitio de unión que es accesible al Ca2+ desde el citoplasma, y se activa mediante fosforilación, usando el Pi derivado del ATP. La Ca2+ ATPasa funciona de forma parecida a la Na+ /K+ ATPasa, con los estados E1 y E2 que tienen afinidades alta y baja por el Ca2+ , respectivamente. La bomba actúa de la siguiente amnera: 1) la unión de un Ca2+ citoplasmático a un sitio aminoácido en la bomba, activa la ATPasa, lo que causa la hidrólisis del ATP hacia ADP y Pi, que están unidos a la bomba; 2) las dos salidas para el Ca2+ ahora están momentáneamente bloqueadas; 3) el ADP se libera, lo que produce un cambio de forma en la proteína, que abre un pasaje para el Ca2+ hacia el líquido extracelular (o cisterna del retículo endoplásmico), de modo que el Ca2+ puede moverse hacia el otro lado de la membrana; 4) el grupo Pi se libera de la bomba, lo que permite que el transportador regrese a su estado inicial donde el Ca2+ citoplasmático de nuevo tiene acceso al sitio de unión.
- 22. BIOLOGIA CELULAR 22 Seminario 1: Transporte a través de la Membrana 2020 – 20 Para la PMCA, el estado E1 fija Ca2+ en el lado intracelular, se produce un cambio conformacional al estado E2, y el estado E2 libera Ca2+ al líquido extracelular. Para la SERCA, el estado E1 fija Ca2+ en el lado intracelular y el estado E2 libera Ca2+ a la luz del retículo sarcoplásmico. La H+ /K+ ATPasa se encuentra en las células parietales de la mucosa gástrica y en las células a- intercaladas del túbulo colector renal. En el estómago, bombea H+ del LIC de las células parietales a la luz del estómago, donde acidifica el contenido gástrico. Esta es la base para secretar ácido clorhídrico en las secreciones digestivas del estómago. En el extremo secretor de las células parietales de las glándulas gástricas la concentración del ion hidrógeno aumenta hasta un millón de veces y después se libera hacia el estómago junto con iones cloruro para formar ácido clorhídrico. 13. Qué características tienen las bombas de tipo V, F y ABC ?. De ejemplos de cada una de ellas. a. Las ATPasas de tipo V. (V de «vacuola ») bombean protones hacia el interior de ciertos orgánulos, como vacuolas, vesículas, lisosomas, endosomas y el complejo de Golgi. La diferencia en la concentración de protones a ambos lados de la membrana de estos organelos, oscila entre 10 y 10.000 veces. Estas ATPasas no se fosforilan durante el transporte. Están formadas por dos componentes multiméricos, uno integral de membrana y otro periférico, que asoma hacia el citosol. Éste último contiene el lugar de unión del ATP y, por tanto, la actividad ATPásica. Figura 10 Proteínas transportadoras de tipo V, F y ABC. b. Las ATPasas de tipo F. (F de «factor») aparecen en bacterias, mitocondrias y cloroplastos. Están implicadas en el transporte de protones y tienen dos componentes multiméricos. El componente integral, llamado Fo es un poro transmembrana de protones. El componente periférico, llamado F1, incluye el sitio de unión al ATP. Las ATPasas de tipo F utilizan la energía de la hidrólisis del ATP, para bombear protones contra su potencial electroquímico. Estas proteínas de transporte pueden facilitar también el proceso contrario, durante el cual, el flujo exergónico de protones a favor de gradiente, se utiliza para sintetizar ATP. Cuando funcionan en este último modo, reciben el término más apropiado de ATP sintasas. Ésta es la forma habitual de funcionamiento de estas proteínas en el metabolismo de energía celular, en el cual, la energía de la radiación solar o de reacciones oxidativas exergónicas, se usa para mantener un gradiente transmembrana de protones, que después servirá para generar ATP. Las ATPasas de tipo F confirman un principio energético importante: El ATP no es sólo la fuente energética necesaria para generar y mantener los gradientes iónicos, sino que estos gradientes pueden servir como fuente de energía para obtener ATP. Este principio, que fue descubierto en los
- 23. BIOLOGIA CELULAR 23 Seminario 1: Transporte a través de la Membrana 2020 – 20 estudios de las ATPasas de tipo de F, es la base de la síntesis de ATP en todos los organismos eucariotas y en la mayoría de los procariotas. c. Las ATPasas de tipo ABC. La designación ABC deriva de «ATP Binding Cassette», donde el término casette hace referencia a los dominios catalíticos de la proteína que unen ATP durante el transporte. Las ATPasas de tipo ABC forman una superfamilia de proteínas transportadoras relacionadas entre sí por secuencias similares y, probablemente, por el mecanismo molecular. Las ATPasas de tipo ABC tienen cuatro dominios, dos de ellos son proteínas integrales de membrana, altamente hidrófobas, mientras que los otros dos son proteínas periféricas, localizadas en la cara citoplásmica de la membrana. Cada uno de los dos dominios integrales consta de seis segmentos transmembrana, que probablemente forman el elemento de paso de las moléculas de soluto. Los dos dominios periféricos son los casettes que unen ATP y acoplan su hidrólisis al proceso del transporte. Las ATPasas de tipo ABC son específicas para un determinado soluto o un grupo de solutos, estrechamente relacionados entre sí. Tomadas en su conjunto, la variedad de solutos transportados por los numerosos miembros de esta superfamilia es grande, incluyendo iones metálicos, azúcares, aminoácidos, péptidos, así como sales biliares, medicamentos y muchas moléculas hidrófobas que no serían capaces de atravesar la bicapa lipídica de otro modo, o lo harían muy lentamente. MDR (del inglés multidrug-resistance). Son transportadores que confieren a las células resistencia frente a múltiples fármacos. Estos transportadores expulsan xenobióticos, antibióticos y otros tipos de fármacos fuera de la célula. Muchas células tumorales aumentan sus niveles de MDR para poder defenderse de los fármacos antitumorales con lo que se trata a los pacientes, lo que constituye un fenómeno de resistencia muy importante para la actuación de estos tratamientos. MDR1, también llamada P-glucoproteína 1, está ampliamente distribuida en el epitelio intestinal, los hepatocitos, epitelio del túbulo contorneado proximal, glándula suprarrenal, etc. Esta proteína es una bomba que utiliza ATP para expulsar un amplio espectro de compuestos xenobióticos. MDR2 se expresa en órganos excretores como el riñón y el hígado, donde excreta conjugados que se obtienen al neutralizar productos tóxicos con sustancias como el ácido glucorónico, el sulfato, el acetato o el glutation, de manera que sean más fáciles de excretar. CFTR. (regulador de la conductancia transmembrana en la fibrosis quística). El gen CFTR codifica para una proteína de canal que permite el paso del Cl- , el cual posee una función muy importante en la creación de la mucosidad respiratoria, el jugo digestivo y el sudor. Esta proteína posee dos dominios con capacidad de hidrolizar el ATP, lo que posibilita la actividad del canal. Cuando esta proteína resulta inactivada por mutaciones, se desarrolla una enfermedad llamada fibrosis quística; en ella, los pacientes no tienen capacidad de expulsar iones Cl- . En los pacientes con fibrosis quística las mutaciones de CFTR pueden afectar de modo diverso a la proteína, pero es frecuente que ésta quede retenida en el retículo endoplásmico, sin llegar a la membrana luminal de las células. 14. Qué tipos de transporte activo secundario existen?. De algunos ejemplos. En el transporte activo secundario (o transporte acoplado) la energía necesaria para el movimiento “cuesta arriba” de una molécula o ion se obtiene a partir del transporte “cuesta abajo” de Na+ hacia la célula. La hidrolisis de ATP mediante la acción de las bombas de Na+ /K+ se requiere de manera indirecta, a fin de mantener concentraciones intracelulares bajas de Na+ . Existen dos tipos de transporte activo secundario y se distinguen por la dirección del movimiento del soluto ascendente. Si el soluto ascendente se mueve en la misma dirección que el Na+ , se llama cotransporte o simporte. Si el soluto ascendente se mueve en la dirección contraria del Na+ , se llama contratransporte, antiporte o intercambio.
- 24. BIOLOGIA CELULAR 24 Seminario 1: Transporte a través de la Membrana 2020 – 20 El cotransporte (simporte) es una forma de transporte activo secundario en el que todos los solutos son transportados en la misma dirección a través de la membrana celular. El Na+ se mueve hacia el interior de la célula en el transportador a favor de su gradiente electroquímico; los solutos, cotransportados con el Na+ , también se mueven hacia el interior de la célula, pero en contra de su gradiente de concentración. El gradiente del ion sodio se mantiene a través de la bomba de Na+ /K+ . Así pues, los iones sodio circulan a través de la membrana plasmática, siendo bombeados hacia exterior por la bomba de Na+ /K+ y regresando a la célula, a la vez que ceden energía para el simporte de moléculas. El cotransporte es característico de varios procesos fisiológicos muy importantes, especialmente en el epitelio absorbente del intestino delgado y el túbulo renal. Por ejemplo, el cotransporte de Na+ /glucosa y el cotransporte de Na+ /aminoácidos se encuentran en las membranas luminales de las células epiteliales del intestino delgado y del túbulo proximal renal. Otro ejemplo en túbulo renal es el cotransporte de Na+ /K+ /2Cl– , que se encuentra en la membrana luminal de las células epiteliales de la rama gruesa ascendente del asa. En cada ejemplo, el gradiente de Na+ establecido por la Na+ /K+ ATPasa se usa para transportar solutos como glucosa, aminoácidos, K+ o Cl– en contra de sus gradientes. El cotransporte de Na+ /glucosa (SGLT1 o proteína transportadora de Na+ /glucosa) en las células epiteliales intestinales. El cotransportador se encuentra en la membrana luminal de estas células y se aprecian dos sitios de reconocimiento específicos, uno para los iones Na+ y otro para la glucosa. Cuando se encuentran Na+ y glucosa en la luz del intestino delgado, se unen al transportador. El primer paso en este proceso es la unión del Na+ extracelular a su sitio de unión con carga negativa sobre la proteína transportadora. Esto permite que la glucosa extracelular se una con afinidad alta a su sitio de unión sobre el transportador. Para una forma del transportador común en el intestino delgado, la proporción es de dos Na+ a una glucosa, en cambio en el transportador de los riñones hay una proporción de un Na+ a una glucosa. El transportador entonces sufre un cambio conformacional (de forma) que transporta el Na+ y la glucosa al interior de la célula (Posteriormente, los 2 solutos son transportados al exterior de la célula a través de la membrana basolateral: el Na+ , por la Na+ /K+ ATPasa y la glucosa, por difusión facilitada.). Después de que el Na+ y la glucosa se liberan, el transportador regresa a su conformación original. Figura 11 Modelo de mecanismo de simporte Na+/glucosa.
- 25. BIOLOGIA CELULAR 25 Seminario 1: Transporte a través de la Membrana 2020 – 20 El cotransporte con sodio de los aminoácidos se produce de la misma manera que para la glucosa, excepto porque utiliza un grupo diferente de proteínas transportadoras. Se han identificado cinco proteínas transportadoras de aminoácidos, cada una de las cuales es responsable de transportar un grupo de aminoácidos con características moleculares específicas. La inhibición de la Na+ /K+ ATPasa reduce el transporte de Na+ del LIC al LEC, lo que produce un aumento de la concentración intracelular de Na+ y, por tanto, el descenso del tamaño del gradiente transmembrana de Na+ . Por tanto, y de forma indirecta, todos los proce- sos de transporte activo secundario están reducidos por los inhibidores de la Na+ /K+ ATPasa porque su fuente de energía (gradiente de Na+ ) está disminuida. En el contratransporte, los iones sodio intentan una vez más difundir hacia el interior de la célula debido a su gran gradiente de concentración. Sin embargo, esta vez la sustancia que se va a transportar está en el interior de la célula y se debe transportar hacia el exterior. Por tanto, el ion Na+ se une a la proteína transportadora en el punto en el que se proyecta hacia la superficie exterior de la membrana, mientras que la sustancia que se va a contratransportar se une a la proyección interior de la proteína transportadora. Una vez que ambos se han unido se produce un cambio conformacional moviéndose el Na+ hacia el interior permitiendo que la otra sustancia se mueva hacia el exterior. Dos mecanismos de contratransporte especialmente importantes son el contratransporte sodio- calcio y el contratransporte sodio-hidrógeno. El contratransporte Na+ /Ca2+ se produce a través de todas o casi todas las membranas celulares, de modo que los iones Na+ se mueven hacia el interior y los iones Ca2+ hacia el exterior, ambos unidos a la misma proteína transportadora. . El intercambio Na+ /Ca2+ es uno de los mecanismos de transporte (además la Ca2+ ATPasa) que ayuda a mantener la concentración intracelular de Ca2+ en valores muy bajos (≈10– 7 molar). Para conseguir el intercambio Na+ /Ca2+ , debe existir un transporte activo, dado que el Ca2+ sale de la célula en contra de su gradiente electroquímico. La proteína de intercambio tiene sitios de reconocimiento para Ca2+ y Na+ . La proteína debe unir Ca2+ en el lado intra- Figura 11 Modelo de mecanismo celular de la membrana y, simultáneamente, Na+ en el lado de antiportador Na+/Ca2+ y Na+/H+. extracelular. En esta configuración, la proteína de intercambio cambia conformacionalmente y libera Ca2+ al exterior de la célula y Na+ a su interior. Generalmente, entran 3 iones Na+ en la célula por cada ion Ca2+ extraído de la misma. Al entrar tres cargas positivas en la célula que se intercambian por dos cargas positivas que salen, haciendo que el intercambiador Na+ /Ca2+ sea electrogénico. El contratransporte Na+ /H+ se produce en varios tejidos. Un ejemplo especialmente importante se produce en los túbulos proximales de los riñones, en los que los iones sodio se desplazan desde la luz del túbulo hacia el interior de la célula tubular, mientras que los iones hidrógeno son contratransportados hacia la luz tubular. Como mecanismo para concentrar los iones hidrógeno, el contratransporte no es en modo alguno tan eficaz como el transporte activo primario de los iones hidrógeno que se produce en los túbulos renales más distales, aunque puede transportar cantidades muy grandes de iones hidrógeno, lo que hace que sea clave para el control del ion hidrógeno en los líquidos corporales.
- 26. BIOLOGIA CELULAR 26 Seminario 1: Transporte a través de la Membrana 2020 – 20 Tabla 4. Ejemplos de cotransportadores y contratransportadores Proteína Sustrato Función fisiológica Cotransportadores (simportadores) Na+ /glucosa (SGLT1) 2 Na+ hacia dentro : 1 glucosa hacia dentro Captación de glucosa en el intestino y resorción por los riñones Na+ /Cl- 1 Na+ hacia dentro : 1 Cl- hacia dentro Resorción de NaCl en los riñones Na+ /K+ /Cl- 1 Na+ hacia dentro : 1 K+ hacia dentro : 2 Cl- hacia dentro Resorción de NaCl en los riñones Na+ /HCO3- 1 Na+ hacia dentro : 1 Cl- hacia dentro Control del pH de líquidos digestivos pancreáticos Na+ /yoduro- 2 Na+ hacia dentro : 1 yoduro hacia dentro Captación de yoduro en la glándula tiroides y otros tejidos Na+ /prolina- 1 Na+ hacia dentro : 1 prolina hacia dentro Captación de prolina en bacterias; transportadores similares para resor-ción de aminoácidos por los riñones Contratransportadores (Intercambiadores / antiportadores) Na+/Ca2+ 3 Na+ hacia dentro : 1 Ca2+ hacia afuera Eliminación de Ca2+ citosólico Na+ /K+ /Ca2+ 4 Na+ hacia dentro : 1 K+ hacia afuera : 1 Ca2+ hacia afuera Adaptación a la luz Na+/H+ 1 Na+ hacia dentro : 1 H+ hacia afuera Regulación del pH intracelular y del volumen celular Na+ /Mg2+ 2 Na+ hacia dentro : 1 Mg2+ hacia afuera Mantenimiento de la concentración intracelular de Mg2+ 15. Cómo se produce el transporte epitelial ? En muchas localizaciones del cuerpo se deben transportar sustancias a través de todo el espesor de una capa celular en lugar de simplemente a través de la membrana celular. El transporte de este tipo se produce a través de: 1) el epitelio intestinal; 2) el epitelio de los túbulos renales; 3) el epitelio de todas las glándulas exocrinas; 4) el epitelio de la vesícula biliar, y 5) la membrana del plexo coroideo del cerebro y otras membranas. El mecanismo básico para el transporte de una sustancia a través de una lámina celular es: 1) transporte activo a través de la membrana celular de un polo de las células transportadoras de la capa, y después 2) difusión simple o difusión facilitada a través de la membrana del polo opuesto de la célula. Las membranas epiteliales cubren todas las superficies del cuerpo y revisten las cavidades de todos los órganos huecos; por ende, para que una molécula o ion se mueva desde el ambiente externo hacia la sangre (y desde ahí hacia los órganos del cuerpo), debe pasar primero a través de una membrana epitelial. El transporte de productos de la digestión (como glucosa) a través del epitelio intestinal hacia la sangre se llama absorción. El transporte de moléculas hacia afuera del filtrado urinario (originalmente derivado de la sangre) de regreso hacia la sangre se llama resorción. El cotransporte de Na+ y glucosa descrito en la sección anterior puede servir como un ejemplo. Los transportadores de cotransporte para Na+ y glucosa están ubicados en la membrana plasmática apical (superior) de las células epiteliales, que mira hacia la luz del intestino o del túbulo renal. Las bombas de Na+ /K+ y los transportadores para la difusión facilitada de glucosa, están en el lado opuesto de la célula epitelial (mirando hacia la ubicación de los capilares sanguíneos). Como resultado de estos procesos de transporte activo y pasivo, la glucosa se mueve desde la luz, a través de la célula, y después hacia la sangre. Los aminoácidos se transportan de manera similar a través del revestimiento epitelial del intestino delgado y de los túbulos renales. Algunos aminoácidos se cotransportan mediante un transportador que utiliza el gradiente electroquímico de Na+ , de manera similar al cotransporte de glucosa; sin embargo, otros aminoácidos son transportados mediante un transportador que utiliza un gradiente electroquímico de protones (H+ ).
- 27. BIOLOGIA CELULAR 27 Seminario 1: Transporte a través de la Membrana 2020 – 20 Este gradiente de H+ se origina por medio de un transportador diferente, una bomba de Na+ /H+ , que usa el movimiento hacia adentro de Na+ para transportar H+ hacia afuera de la célula. Los mecanismos de transporte de membrana descritos en esta sección mueven materiales a través del citoplasma de las células epiteliales, proceso denominado transporte transcelular. Empero, la difusión y osmosis también pueden ocurrir a un grado limitado en los muy pequeños espacios entre las células epiteliales, proceso conocido como transporte paracelular. El transporte paracelular entre células está limitado por los complejos de unión los cuales conectan células epiteliales adyacentes. Figura 12 Transporte transcelular de glucosa desde la luz intestinal hacia la sangre El grado al cual los complejos de unión rodean cada célula epitelial determinara cuanto transporte paracelular será posible entre ellas. Por ejemplo, las células epiteliales que componen las paredes de muchos capilares sanguíneos (los vasos sanguíneos de calibre más delgado) tienen poros entre ellos que pueden ser relativamente grandes, lo que permite la filtración de agua y moléculas disueltas hacia afuera de los capilares a través de la ruta paracelular. Con todo, en los capilares del cerebro, esa filtración se evita mediante zonas de oclusión, de modo que las moléculas deben transportarse de manera transcelular.
- 28. BIOLOGIA CELULAR 28 Seminario 1: Transporte a través de la Membrana 2020 – 20 2. BIBLIOGRAFÍA ALBERTS B., et al. Biología Molecular de la Célula. 5ª ed. Cápitulo 11: Transporte de pequeñas moléculas a trravés de la membrana y las propiedades eléctricas de las membranas. Editorial Médica Panamericana S.A. 2010: 1728p. ALBERTS B. et al. Introducción a la Biología Celular. 3ª ed. Cápitulo 12: Transporte de Membrana Editorial Médica Panamericana S.A. 2011: 900p. BARRET K.E. et al. Ganong Fisiología Médica. 24ª ed. Capítulo 6: Transporte de iones y moléculas pequeñas a través de la membrana. Editorial Mc Graw-Hill, 2011: 1054p BECKER W. HARDIN J. KLEINSMITH L. El Mundo de la Célula. Capítulo 8: Transporte a través de membrana: saltando la barrera de permeabilidad. 6ª ed. Editorial Pearson Prentice Hall. 2007. CALVO A. Biología Celular Biomédica. 1° ed. Cápitulo 4: Microtransporte a través de la Membrana Plasmática. Editorial Elsevier Science. 2015. CASSIMERIS L., LINGAPPA V.R., PLOPPER G. Células. 2ª ed. Capítulo 6: Transporte de iones y moléculas pequeñas a través de la membrana. Editorial Mc Graw-Hill, 2011. CHANDAR N., VISELLI S. Biología Molecular y Celular. 1° ed. Cápitulo 13: Conceptos Básicos de Transporte, Cap. 14: Transporte Activo, Cap.15: Transporte de glucosa. Editorial Lippincot Williams & Wilkins. 2011 CONSTANZO. L. Fisiología. 5° ed. Capítulo 1: Fisiología Celular. Editorial Elsevier Saunders. 2014. COOPER G. La Célula. 5° ed. Cápitulo 13: Membrana Plasmática. Edit. Marbán. 2010. DE ROBERTIS E.M.F., HIB J. Fundamentos de Biología Celular y Molecular. 4ª ed. Cápitulo 3: Las Membranas Celulares. Buenos Aires: El Ateneo, 2004: 442p. FOX. S.I. Fisiología Humana 12 ed. Capítulo 6: Interacciones entre Células y el Ambiente Extracelular. México: Editorial Mc Graw-Hill, 2011: 749p. HALL. J. Tratado de Fisiología Médica. 11° ed. Transporte de Sustancias a Través de las Membranas Celulares. Barcelona: Editorial Elsevier 2011:1092p. KARP G. Biología Celular y Molecular. Conceptos y Experimentos. 6ª ed. Cápitulo 8: Sistemas de la Membrana Citoplásmica: estructura, función y tráfico de membrana. México: Editorial Mc Graw-Hill, 2011: 899p. LODISH H., et. al. Biología Celular y Molecular. 5ª. ed. Cápitulo 7: Transporte de Iones y Pequeñas Moléculas a través de las Membranas Celulares. Madrid: Editorial Médica Panamericana S.A., 2005: 1030p. SILVERTHORN. Fisiología Humana. Un enfoque Integrado. 4ª ed. Cápitulo 5: Dinámica de las Membranas. Editorial Médica Panamericana S.A. 2007. STANFIELD C.L: principios de Fisiología Humana. Capítulo 4: Transporte de la Membrana Celular. 4ª ed. Editorial Pearson Educación. 2011.