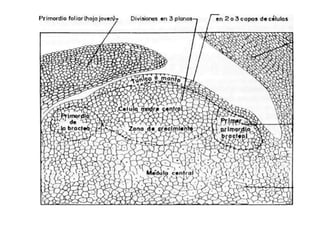
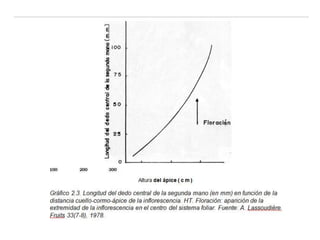
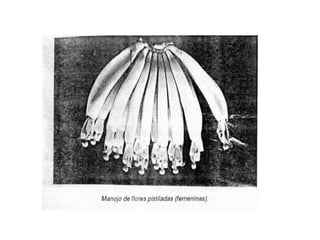
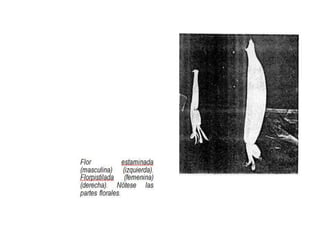
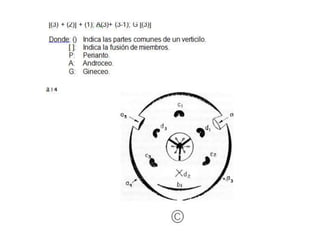
La inflorescencia del banano se desarrolla a partir de una yema floral en el punto de crecimiento de la planta. La inflorescencia crece hacia arriba a través del centro del pseudotallo durante más de 100 días hasta emerger. Contiene flores masculinas y femeninas agrupadas en manojos, con las flores femeninas diferenciándose primero en pequeños números antes de grandes cantidades de flores masculinas. El desarrollo de la inflorescencia depende de factores como el estado nutricional y de crecimiento de la planta.