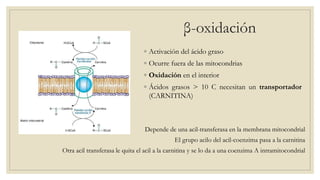
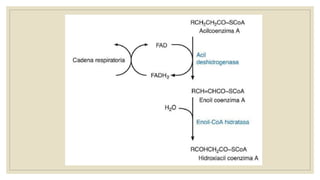
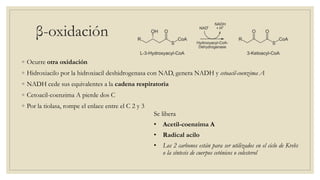
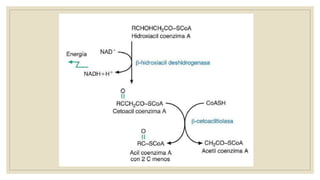
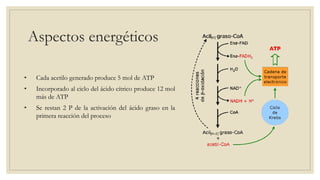
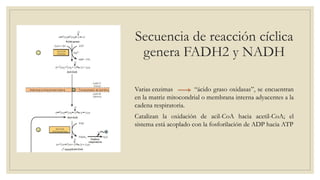
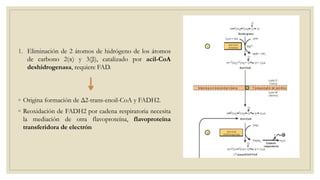
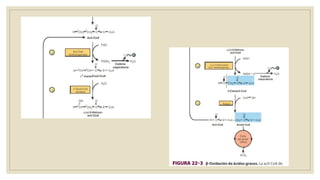
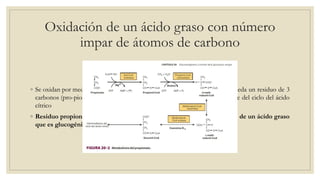
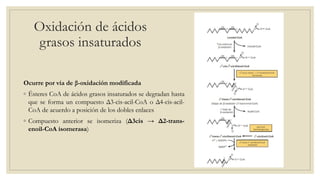
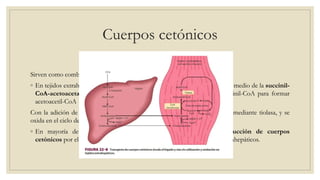
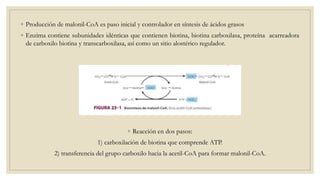
Los ácidos grasos libres se activan mediante la formación de acil-CoA antes de ser catabolizados a través de la β-oxidación. La β-oxidación consiste en la oxidación secuencial de acil-CoA para liberar moléculas de acetil-CoA de dos carbonos, generando energía en forma de FADH2 y NADH. Los ácidos grasos de cadena larga requieren ser transportados al interior de la mitocondria mediante la carnitina para ser degradados.