Descargado 13 veces


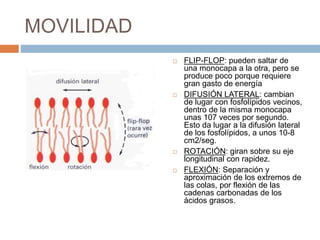

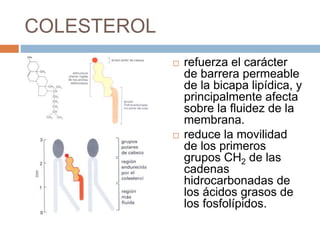
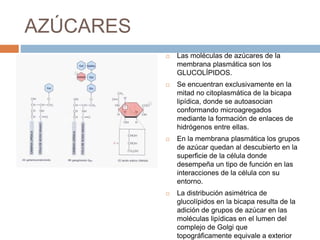


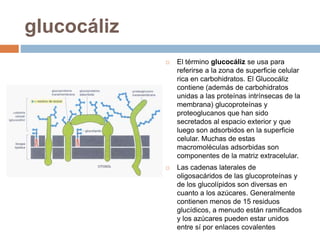
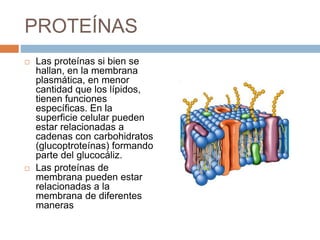
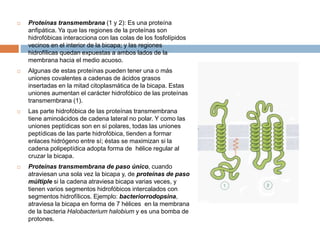



Este documento describe la composición de las membranas biológicas. Las membranas están compuestas principalmente por lípidos como fosfolípidos y colesterol. También contienen proteínas y azúcares unidos a lípidos y proteínas. Las proteínas pueden ser transmembrana o periféricas y tienen funciones importantes como el transporte a través de la membrana. Los azúcares forman parte del glucocáliz que recubre la superficie celular.

























































![CAP 11 ESTRUCTURA DE MEMBRANA [Autoguardado].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/cap11estructurademembranaautoguardado-260119035255-d0910a3c-thumbnail.jpg?width=640&height=640&fit=bounds)

















































