Descargado 28 veces












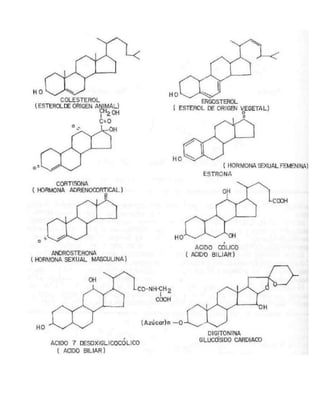













































































Este documento trata sobre los lípidos, sus estructuras y funciones. Explica que los lípidos son compuestos heterogéneos que comparten propiedades como su insolubilidad en agua y solubilidad en solventes no polares. Describe que cumplen funciones estructurales como componentes de membranas y funciones energéticas al almacenar calorías. También habla sobre diferentes tipos de lípidos como fosfolípidos, esfingolípidos, glicolípidos y sus roles en el cuerpo.























































