
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (20)
Similar a Membrana plasmatica
Similar a Membrana plasmatica (20)
Último
Último (20)
Membrana plasmatica
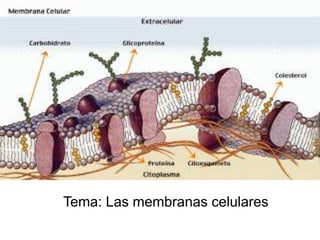
- 1. Tema: Las membranas celulares
- 2. Las membranas celulares ejercen diversas actividades _ la célula se halla rodeada por la membrana plasmática, una delgada capa de 6 a 10 nm de espesor compuesta por lípidos proteínas e hidratos de carbono . _Su estructura básica es similar alas de las restantes membranas de la célula las cuales envuelven alos organoides del sistema de endomenbranas incluida en la envoltura nuclear. _ las membranas celulares ejercen actividades complejas como son las siguientes.
- 3. Constituyen verdaderas barreras permeables selectivas que controlan el pasaje de iones y de moléculas pequeñas impide el intercambio de los componentes de los organoides entre si y de los componentes extracelulares de la célula. Proveen soporte físico para la actividad ordenada de las enzimas que se asientan en ellas. Mediante la formación de pequeñas vesículas transportadoras hacen posible el desplazamiento por el citoplasma La membrana plasmática participa en los procesos de endocitosis y de exocitosis. En la membrana plasmática existen moléculas mediante las cuales las células se reconocen y se adhieren entre si.
- 6. las soluciones acuosas puras de los fosfolípidos no forman mono capas sino bicapas que sierran sobre las mismas en lo cual da a las vesículas llamadas liposomas
- 7. Cuando se coloca fosfolípidos entre las soluciones acuosas separadas por un tabique implementado forman una bicapa lapida que completa la separación
- 8. los enlaces dobles de las cadenas saturadas no producen angulosidades en los ácidos grasos lo cual separan alos fosfolípidos y le da ala bicapa una configuración menos compacta
- 9. El colesterol es un componente cuantitativamente importante de las membranas celulares especialmente en las membranas plasmáticas debido a que es antipático en cada mono capa
- 10. La bicapa lápida se comporta como una estructura fluida , la fluidez aumenta significa que sus compontes rotan entorno a sus ejes y se desplazan firmemente por la superficie membranosa además estos movimientos pueden pasar por una capa ala otra por un movimiento llamado flipflop .
- 11. La fluidez de las proteínas en la bicapa lipidia ha suido comprobada mediante distintas técnicas biológicas Mayor información otorga el uso de anticuerpos ligados a fluorocromos que son fáciles de detectar mediante el microscopio de fluorescencia y mediante este proceso se puede observar el desarrollo de una especie de capuchón.
- 12. Si previamente se marcan las células con sendos anticuerpos fluorescentes de colores diferentes luego de la fusión pueden reconocerse en la membrana plasmática del heterocarion las partes aportadas por cada célula
- 13. • Se representa el posible mecanismo molecular de fusión de membranas en donde suceden los siguientes fenómenos • Se despejan las proteínas membranosas dejando a las bicapas sin otro tipo de moléculas que los lípidos. • Las bicapas establecen íntimo contacto a través de sus respectivas mono capas enfrentadas. • Dichas capas desaparecen y se desarrolla una interfase de estructuras lipidicas hexagonales. • Finalmente la interfase desaparece y se completa la fusión.
- 14. Los hidratos de carbono de las membranas celulares forman parte de glicolípidos y glicoproteínas • Cerebrocidos • Gangleocidos • Glicoproteínas Los oligosacáridos se hallan ligados a proteínas mediante enlaces N-glicosidicos, una proteína puede contener una o varias cadenas oligosacáridos.
- 15. Los hidratos de carbono cumplen funciones relevantes en las membranas celulares • Protegen a la superficie de la célula de agresiones químicas y mecánicas (células de la mucosa intestinal que son protegidas del contacto con los alimentos). • Por la presencia de ácidos sialicos en oligosacáridos la carga eléctrica en su superficie es negativa, esta condición es importante en células nerviosas y musculares.
- 16. • En algunas capilares sanguíneos el glicocaliz se comporta como un filtro que regula el paso de moléculas según sus tamaños y propiedades químicas • Algunos oligosacáridos del glicocaliz son necesarios en los procesos de reconocimiento y adhesión celular. • La especialidad del sistema ABO de grupos sanguíneos es determinada por ciertos oligosacáridos muy cortos y parecidos entre si presentes en los glóbulos rojos. • En células tumorales malignas se han observado cambios en algunos oligosacáridos ya que alteran la recepción de las señales que controlan las divisiones celulares. • En ciertos tejidos que contienen componentes del glicocaliz presentan propiedades enzimáticas.
- 17. PERMEABILIDAD DE LAS MEMBRANAS CELULARES • Los solutos y macromoléculas atraviesan las membranas por mecanismos diferentes. Existe un flujo continuo de sustancias que entran y salen de la célula y circulan por su interior ,para ello los solutos deben pasar a través de la membrana celular a este fenómeno se le denomina permeabilidad celular. En macromoléculas ,para atravesar las membranas , algunos ocupan canales membranosos especiales llamado translacones .Otros pasan por poros de sofisticada composición y otras se valen de pequeñas vesículas.
- 18. EL PASAJE DE SOLUTO A TRAVES DE LAS MEMBRANAS CELULARES PUEDE SER PASIVO O ACTIVO • El incesante intercambio de solutos entre la célula y el medio que la rodea . Según el tipo de soluto el paso se produce con energía o por mecanismos que requieren energía. • Cuando no consume energía se denomina transporte pasivo y el dependiente de energía transporte activo.
- 19. TRANSPORTE PASIVO Se cumple a través de los componentes de la bicapa lipídica Por proteínas transmembranosas para el paso de solutos Estas estructuras son de 2 tipos. Canales iónicos Permeasas El transporte pasivo a través de la bicapa lipídica se denomina difusión simple y cuando se realiza por canales iónicos se la llama difusión facilitada . El transporte activo tiene lugar exclusivamente a través de permeasas.
- 20. EL TRANSPORTE PASIVO DE SOLUTOS SE PRODUCE POR DIFUSION Cuando se disuelve un soluto en un solvente , las partículas del primero se dispersan hasta quedar uniformemente distribuidas. La difusión o movimiento de un soluto se da de los sitios de menor concentración a los de mayor concentración La difusión a favor del gradiente de concentración , esta ocurre sin gasto de energía de ahí en nombre de transporte pasivo
- 21. TRANSPORTE PASIVO Se cumple a través de los componentes de la bicapa lipídica Por proteínas transmembranosas para el paso de solutos Estas estructuras son de 2 tipos. Canales iónicos Permeasas El transporte pasivo a través de la bicapa lipídica se denomina difusión simple y cuando se realiza por canales iónicos se la llama difusión facilitada
- 22. La difusión simple se produce a través de la bicapa lipídica • El transporte pasivo puede ocurrir también entre compartimientos acuosos separados por membranas semipermeables. Como lo son las bicapa lipídica de las membranas . • A este tipo de transporte se lo denomina difusión simple. • Las sustancias que se disuelven en los lípidos atraviesan con cierta facilidad la zona hidrofobica.
- 23. La difusión facilitada se produce a través de canales iónicos y permeasas • La mayoría de sustancias que atraviesan las membranas a favor del gradiente de concentración lo hacen a una mayor velocidad esperable si su pasaje fuera por difusión simple. La diferencia se explica por la presencia de ciertos componentes membranosos proteicos llamados canales iónicos y permeasas , a través de los cuales se facilita aunque también se regula la transferencia de los solutos de un lado al otro de la membrana.
- 24. • Como vemos en la difusión facilitada la fuerza que impulsa la movilización de las partículas del solutos es el gradiente , y por lo tanto no consume energía . • Desde este punto de vista la difusión facilitada es similar ala difusión simple ; la diferencia reside en que en la primera actúan estructuras proteicas reguladoras y en la segunda no .
- 25. CANALES IONICOS Los canales iónicos son poros o túneles hidrofilicos que atraviesan las membranas ,formados por proteínas integrales transmembranosas generalmente de tipo multipaso Son altamente selectivos , de modo que ay canales específicos para cada tipo de ion (Na+,k+,Ca2+, etc.)Los mas abundantes en la membrana son los canales para el K+ Clases de canales iónicos DEPENDIENTES DEL VOTAJEDEPENDIENTES DEL LIGANDO
- 26. La mayoría de los canales iónicos no están abiertos en forma permanente , pues poseen un dispositivo de apertura y cierre de alguna compuerta a respuesta a un cambio en el potencial eléctrico a estos se le denomina dependientes del voltaje , y otros abren el mecanismos cuando le llega una sustancia inductora por el lado citosólico o no citosólico a estos canales se los denomina dependientes de ligando.
- 27. LOS IONOFOROS AUMENTAN LA PERMEABILIDAD DE LAS MEMBRANAS BILOGICAS A CIERTOS IONES • Existen sustancias llamadas ionóforos que tienen la propiedad de incorporarse alas membranas biológicas y aumentar su permeabilidad con diversos iones
- 28. IONOFOROS Son moléculas de tamaño relativamente pequeño ,con una superficie hidrofobica que les permite insertarse en la bicapa lipídica Clases de ionóforos Formadores de canalesTransportadores móviles Estos atrapan al ion en un lado de la membrana ,lo engloban en el interior de sus moléculas , giran en la bicapa y lo liberan del otro lado. So0n conductos hidrofóbicos el pasaje de cationes monovalentes (H+,Na+,K+)
- 29. ACUOPORINAS Son canales especiales que permiten el paso selectivo del agua Las acuoporinas se hallan constituidas por cuatro proteínas transmembranosas de 28kDa cada una denominadas CHIP (por channel- forming integral protein) cuya estructura secundaria es helicoidal estas se asocian a un canal central , evidencias experimentales demuestran que cada hélice se comporta como un poro individual El pasaje de las moléculas de agua se realiza se realiza sin la compañía de iones ni de otro tipo de solutos .Aun no se han Establecido las bases de esta especificad
- 30. PERMEASAS La pared de las permeasas esta constituida por varias proteínas transmembranosas multipaso. Cada permeasa posee sitios de unión específicos para uno o dos clases de solutos ,accesibles desde ambas caras de la bicapa Existen tres clases de permeasas Contransporte (antiport) Monotransporte(en ingles uniport) Cotransporte (symport ) Se refieren a un solo tipo de soluto Transportan dos tipos de soluto simultáneamente ambos en el mismo sentido . Aquí se hallan acoplados obligadamente es decir una no se produce sin la otra . Se transfieren dos tipos de solutos en sentidos contrarios
- 31. TRANSPORTE ACTIVO REQUIERE ENERGÍA • Cuando el transporte de un soluto se realiza en dirección contraria a su gradiente de concentración o voltaje, solo es posible con gasto de energía (transporte activo). • El transporte activo tiene lugar a través de permeasas y también es este caso existen formas de monotransporte, cotranporte y contratransporte. El transporte activo de solutos presenta las mismas característica de especificidad y saturabilidad señaladas por la difusión facilitada.
- 32. LA BOMBA DE NA Y K ES UN SISTEMA DE CONTRATRANPORTE • la bomba de Na+ K+ ATP asa tiene por función expulsar Na+ al espacio extracelular e introducir K+ al citosol, dado que transfiere diferentes solutos en sentidos contrarios se trata de un sistema de contratransporte. • Es un complejo integrado por 4 subunidades: 2α y 2β las cuales son proteínas integrales de la membrana plasmática. • La transferencia de Na+ hacia el exterior y K+ hacia el citosol se hallan acopladas, ya que una no puede realizarse sin la otra así que ambos flujos se realizan en contra de sus respectivas gradientes de concentración. • Este sistema necesita de energía que se obtiene por hidrolisis del ATP, por cada ATP que se hidroliza posibilita el transporte de tres Na+ hacia el espacio extracelular y dos K+ hacia el citosol.
- 33. • La bomba de Na+ K+ se la puede interpretar con la siguiente reacción: 3Na + 2K + ATP 3Na + 2K + ADP + P • las bombas q generan potenciales eléctricos de membrana se las define como electrogénicas.
- 34. ALGUNOS FARMACOS CARDIACOS INHIBEN LA BOMBA DE Na K • La digixina y degitoxina son ampliamente utilizados como cardiotónicos, los cuales bloquean el contratransporte de Na y K. estas sustancias actúan en la superficie de las células uniéndose a los sitios de las subunidades α reservados para el K, ya que estas sustancias impiden la liberación del fosfato ligado a la sub unidad α del transportador. • Como consecuencia el sistema se bloquea y disminuye la salida de Na al medio extracelular. DIVERSOS TRANSPORTADORES PASIVOS, AUNQUE AJENOS A LA BOMBA DE Na K FUNCIONAN BAJO SU DEPENDENCIA • La dependencia de contratransporte de Na+ y Ca2+ de la actividad de la bomba de Na K solo es un ejemplo del funcionamiento normal de la celula.
- 35. • En consecuencia, si la bomba de Na K se detiene los transportadores pasivos que dependen de ella dejan de funcionar. • También es el contratransporte de Na+ y H+, el Na ingresa en el citosol y se intercambian por H+ el cual es expulsado de la célula para la regulación del PH de la misma. • Una bomba de K H es responsable de la formación del HCl gástrico. • En las células parietales de la mucosa gástrica existe una bomba de K H cuya estructura no es bien conocida, esta da lugar al contratransporte de K+ y H+ con gasto de energia. • Esto hace que se incrementen los niveles de K+ en el citosol y permiten que alcancen elevadas concentraciones de H+ en la secreción gástrica, esta es acompañada por la salida de Cl-, que en la luz del estómago forman HCl.
- 36. • El K+ y Cl-(proviene de la sangre) salen de la célula por sendas permeasas monotransportadoras. DISTINTAS BOMBAS DE Ca2+ MANTIENEN LA CONCENTRACIÓN DEL ION EN EL CITOSOL EN NIVELES MUY BAJOS • Existen bombas de Ca2+ en la membrana plasmática, membrana del retículo endoplasmática estas poseen sitos de alta afinidad para el Ca2+ en la caracitosólica de ambas membranas, estas bombas de calcio necesitan de Mg2+ y energía del ATP. LA BOMBA DE H+ DISMINUYE EL PH DE LOS LISOSOMAS • Una concentración alta de H+ en el interior de los lisosomas es crucial para la activación de enzimas hidrofílicas las cuales actúan solo cuando el PH es 5.0. • El transporte de H+ desde el citosol al interior del lisosoma depende de la bomba de H+.
- 37. LAS MDR SON TRANSPORTADORES QUE CONFIEREN A LAS CELULAS RESISTENCIA A CIERTAS DROGAS • Son proteínas que pertenecen a una familia de transportadores activos (sus siglas ABC), estas hidrolizan el ATP que provee de energía para mover determinados solutos en contra de sus gradientes. • Los transportadores ABC se encuentran en muchas membranas las cuales tienen la capacidad de eliminar sustancias tóxicas derivadas del metabolismo celular normal. • Estos transportadores MDR cuando aparecen en grandes cantidades en la membrana de células cancerosas contienen resistencia contra algunas drogas citotóxicas, las MDR bombean a estas drogas a fuera de las células cancerosas haciéndolas resistentes a la quimioterapia.
- 38. • Las proteínas MDR ayudan a la resistencia del VIH a drogas antivirales como AZT y así podemos citar muchos mas ejemplos como los parásitos. EN LA FIBROSIS QUISTICA SE HALLA ALTERADO UN CANAL IÓNICO PARA EL Cl2- • Esta se produce por las secreciones muy viscosas que obstruyen la luz de los bronquios, los conductos de varias glándulas (páncreas), tubo intestinal, etc. • La proteína CFTR pertenece a la familia ABC, esta proteína en ciertas células no actúa como permeasa sino como un canal iónico.
- 39. MEMBRANA PLASMATICA Y LA PARED DE LA CELULA VEGETAL LA MEMBRANA PLASMATICA DE UNA CELULA VEGETAL SE HALLA RODEADA POR UNA ESPECIE DE EXOESQUELETO • La gruesa pared celular que recubre a la membrana plasmática es como si fuera un exoesqueleto este le da protección, forma, también mantiene el balance entre la presión osmótica intracelular y la tendencia de penetrar el agua al citosol.
- 40. LA PARED CELULAR CONTIENE UN RETICULO MICROFIBRILAR. • Las microfibrillas de la pared celular están compuestas por celulosa, estas microfibrillas tienen unos 25 nm de diámetro. Algunas paredes celulares pueden contener algunas sustancias cuticulares (ceras) y depósitos minerales como silicatos, carbonatos de sodio y magnesio. LA PARED CELULAR SE COMPONE DE UNA PARED PRIMARIAS Y UNA SECUNDARIA En algunos vegetales podemos encontrar: • Pared primaria • Pared secundaria Las cuales de distinguen por sus matrices y sus microfibrillas. • La pared primaria comienza a formarse con la división celular (telofase) a partir de una placa celular, la misma que esta compuesta por vesículas del complejo de Golgi las cuales forman la capa intermedia de la futura pared celular.
- 41. • Dicha capa solo contiene pectina el cual es un compuesto amorfo. • La célula solo cuando alcanza su madurez aparece la pared secundaria la cual esta formada por celulosa, hemicelulosa y escasas sustancias pécticas.
- 42. ALGUNOS DE LOS COMPONENTES DE LA PARED CELULAR SE ORIGINAN EN EL COMPLEJO DE GOLGI • La intervención del complejo de Golgi es muy evidente ya que ciertas algas cuyas paredes están formadas por escamas, las membranas del complejo de Golgi polimerizan cadenas de glucosa para formar microfibrillas de celulosa por medio del glucosiltransferasas. • Las microfibrillas se organizan y se liberan en la superficie.