Descargado 32 veces




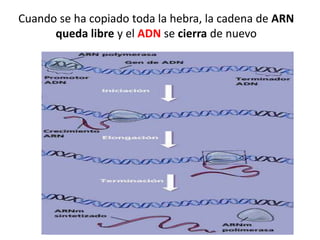




















La transcripción copia la información genética del ADN al ARN. Existen tres tipos de ARN polimerasa en eucariotas que sintetizan diferentes tipos de ARN, mientras que en procariotas y arqueas hay una sola ARN polimerasa. La transcripción implica las etapas de iniciación, elongación y terminación mediante la unión de la ARN polimerasa al promotor y la síntesis del ARN siguiendo la complementariedad de bases del ADN.


![Transcripsion en eucariotas[1]](https://cdn.slidesharecdn.com/ss_thumbnails/transcripsioneneucariotas1-121021194324-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)







































































