
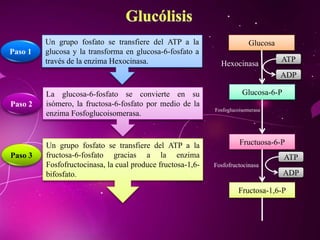
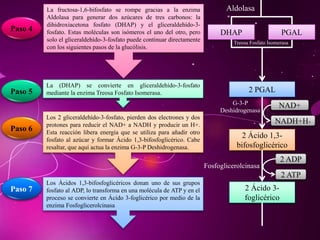
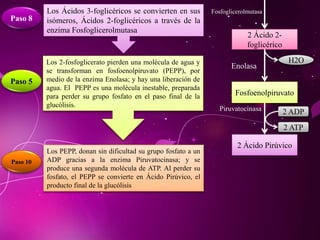
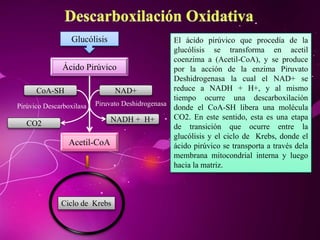



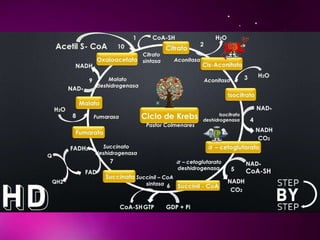

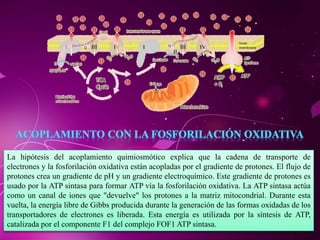
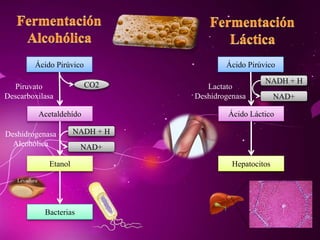
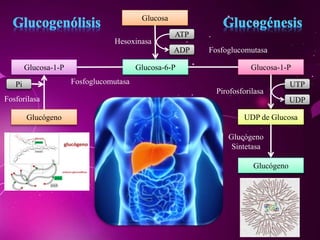


El documento describe los 10 pasos de la glucólisis, en los cuales las enzimas hexocinasa, fosfoglucoisomerasa, fosfofructocinasa, aldolasa, treosa fosfato isomerasa, G-3-P deshidrogenasa, fosfoglicerolcinasa, fosfoglicerolmutasa, enolasa y piruvatocinasa catalizan reacciones que convierten la glucosa en piruvato con la producción neta de ATP y NADH.























































































