Descargado 61 veces























![Cinética enzimática: Efecto de la concentración de S
VR
E+S
E+P
V -R
V1
E+S
V2
ES
V -1
E+P
V -2
V2 << V1 por lo que VR (de la reacción global) depende de V2
Si VR depende de V2, entonces depende de la [ES]
• Si hay poco S, hay poco ES por lo que la reacción será más lenta
• Si hay mucho S, hay mucho ES por lo que la reacción será más rápida
• Si hay exceso de S, toda la E disponible está como ES, por lo que se
alcanza la velocidad máxima de reacción
http://www.biorom.uma.es/contenido/UPV_EHU/enzimas/enz1.htm#p](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-24-320.jpg)




![Ecuación de Michaelis – Menten. Significado de Vmáx
• es una constante.
• Es la velocidad máxima teórica de la
reacción – pero, en realidad, nunca se
alcanza.
• Para alcanzar la velocidad máxima se
requiere que [S] >> [E] y que todas las
moléculas de enzima estén unidas al
sustrato.
• Vmax se alcanza asintóticamente a
medida que la concentración de
sustrato aumenta](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-29-320.jpg)


![Ecuación de Michaelis – Menten: Factores a considerar.
Para utilizar la ecuación de Michaelis-Menten debe asumirse que:
-
Las concentraciones relativas de E y S: La concentración de
sustrato ([S]) es mucho mayor que la concentración de enzima ([E]),
de manera que la proporción de sustrato fijo a la enzima es siempre
relativamente pequeña.
-
La reacción esta en equilibrio: [ES] no cambia durante el tiempo
(la reacción se asume en equilibrio de flujos).
-
Velocidad Inicial: Deben usarse velocidades iniciales (Vo). Esto
significa que la velocidad de la reacción debe determinarse tan
pronto como el sustrato y la enzima son mezclados. En dicho
tiempo, la concentración de productos es despreciable y, por lo
tanto, la reacción inversa de productos a sustratos puede ser
ignorada.](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-32-320.jpg)
![Los dobles recíprocos: Ecuación de Lineweaver-Burk
y=mx+n
y equivale a 1/V0
m equivale a Km/Vmax
x equivale a 1/[S]
n equivale a 1/Vmax](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-33-320.jpg)





![Constante catalítica
k1
E+S
E+S
k1
k-1
k2
ES
k-1
ES
k2
k-2
E+P
EP
k3
kcat = Vmax / [ET]
Vmax = k2·[ET]
k2 = Vmax/[ET]
Vmax = k3·[ET]
k3 = Vmax/[ET]
E+P](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-39-320.jpg)
![Número de recambio
kcat = Vmax / [ET]
NÚMERO DE RECAMBIO
Número de moléculas de sustrato convertidas en
producto en una unidad de tiempo, por una molécula
de enzima cuando la enzima está saturada de
sustrato.](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-40-320.jpg)


![Enzimas alostéricas.
Hay enzimas que no obedecen la ecuación de Michaelis-Menten.
• Se dice que su cinética no es Michaeliana.
• Esto ocurre con las llamadas enzimas alostéricas, cuya gráfica v contra [S] no
es una hipérbola, sino una sigmoide (en forma de s).
• En la cinética sigmoidea, pequeñas variaciones en la [S] en una zona crítica
(cercana a la Km) se traduce en grandes variaciones en la velocidad de
reacción.](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-43-320.jpg)






![Inhibición enzimática: Inhibición competitiva
−
1
Km
Km
aumenta
aumenta
[I]
α = 1+
kI](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-51-320.jpg)

![Inhibición enzimática: Inhibición acompetitiva
[I ]
α ' = 1+
k 'I
1
Vmax
Vmax
−
1
Km
Km
disminuye
disminuye
aumenta
disminuye](https://image.slidesharecdn.com/clase17-enzimas-131214162140-phpapp02/85/Clase-17-enzimas-53-320.jpg)





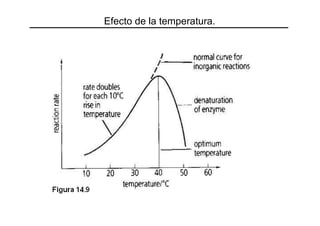
Este documento trata sobre las enzimas y la cinética enzimática. Explica conceptos como la velocidad de reacción, los factores que determinan esta velocidad, las clasificaciones de enzimas, su función y cómo trabajan. También describe la ecuación de Michaelis-Menten, parámetros como Km y Vmax, y diferentes transformaciones gráficas de esta ecuación. Por último, aborda factores que afectan la actividad enzimática como el pH y la temperatura.



































































