Termodinámica y Bioenergética
•Descargar como DOCX, PDF•
1 recomendación•3,331 vistas

Conceptos básicos de termodinámica y bioenergética
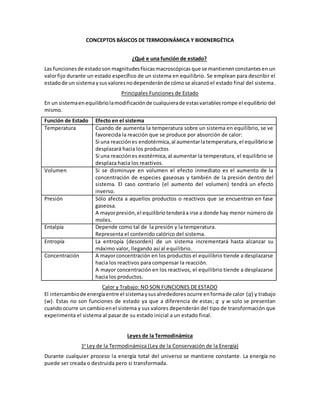
![Representación matemática
ΔE = Q - w
Donde:
ΔE = Cambioenla energíainternadel sistema(Joules,J).
Q= Calor intercambiadoporel sistemaconel entorno (Joules,J).
W= Trabajo intercambiadoporel sistemaconel entorno (Joules,J).
Tambiénse puede expresarcomo:
ΔE = Ef - Ei
Donde
ΔE = Cambioenla energíainternadel sistema.
Ef = Energía internadel sistemaensuestadoinicial.
Ei = Energía internadel sistemaensuestadofinal
2a
Ley de la Termodinámica (Ley de la espontaneidad termodinámica)
El universosiempre tenderáal incrementode suentropía(medidade desordeno aleatoriedad).
Durante cualquierprocesode cambio yase físicoo químico,loselementosdel universotenderána
desordenarse.
Expresión matemática
En un proceso espontáneo: ΔSsistema + ΔSentorno = ΔSuniverso > 0
En un proceso en equilibrio: ΔSsistema + ΔSentorno = ΔSuniverso = 0
Energía libre de Gibbs
La energía libre de Gibbs se define como la cantidad de energía capaz de realizar un trabajo
durante una reacción a condiciones constantes. Puede ser usada como un indicador de la
espontaneidad de una reacción.
Gibbs y su relación con la constante de equilibrio (Keq)
La constante de equilibrio de una reacción, es la expresión de la proporción que hay entre la
concentración de reactantes y la concentración de productos en el punto de equilibrio de la
reacción.
Keq =
[𝑃𝑟𝑜𝑑𝑢𝑐𝑡𝑜𝑠] 𝑒𝑞
[𝑅𝑒𝑎𝑐𝑡𝑖 𝑣 𝑜𝑠] 𝑒𝑞
El ΔG permite calcular que tan alejada se encuentra la reacción del equilibrio bajo condiciones
específicas y que tanta energía se liberara conforme la reacción proceda hacia al equilibrio, por
ejemplo:
En la reacción A B para calcularΔG usandola concentracione prevalentede reactantes(A) y
productos (B) se utiliza la ecuación:
ΔG=RTln
[𝐵] 𝑝𝑟
[𝐴] 𝑝𝑟
- RTln
[𝐵] 𝑒𝑞
[𝐴] 𝑒𝑞
= RTln
[𝐵] 𝑝𝑟
[𝐴] 𝑝𝑟
– RTln Keq
Donde:
ΔG= Cambio de energía libre (cal/mol).
R=Constante de los gases ideales (1.987cal/mol.K).
T=Temperatura (K).
[𝐴] 𝑝𝑟= Concentración prevalente de reactivos (moles/litro, M).](data:image/gif;base64,R0lGODlhAQABAIAAAAAAAP///yH5BAEAAAAALAAAAAABAAEAAAIBRAA7)
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (20)
Similar a Termodinámica y Bioenergética
Similar a Termodinámica y Bioenergética (20)
Más de Campos V
Último
Último (20)
Termodinámica y Bioenergética
- 1. CONCEPTOS BÁSICOS DE TERMODINÁMICA Y BIOENERGÉTICA ¿Qué e una función de estado? Las funcionesde estadoson magnitudesfísicasmacroscópicas que se mantienenconstantesen un valorfijo durante un estado específico de un sistema en equilibrio. Se emplean para describir el estadode un sistemaysusvaloresnodependeránde cómose alcanzóel estado final del sistema. Principales Funciones de Estado En un sistemaenequilibriolamodificaciónde cualquierade estasvariablesrompe el equilibrio del mismo. Función de Estado Efecto en el sistema Temperatura Cuando de aumenta la temperatura sobre un sistema en equilibrio, se ve favorecida la reacción que se produce por absorción de calor: Si una reacciónes endotérmica,al aumentarlatemperatura, el equilibriose desplazará hacia los productos Si una reacciónes exotérmica,al aumentar la temperatura, el equilibrio se desplaza hacia los reactivos. Volumen Si se disminuye en volumen el efecto inmediato es el aumento de la concentración de especies gaseosas y también de la presión dentro del sistema. El caso contrario (el aumento del volumen) tendrá un efecto inverso. Presión Sólo afecta a aquellos productos o reactivos que se encuentran en fase gaseosa. A mayorpresión,el equilibriotenderáa irse a donde hay menor número de moles. Entalpía Depende como tal de la presión y la temperatura. Representa el contenido calórico del sistema. Entropía La entropía (desorden) de un sistema incrementará hasta alcanzar su máximo valor, llegando así al equilibrio. Concentración A mayorconcentración en los productos el equilibrio tiende a desplazarse hacia los reactivos para compensar la reacción. A mayor concentración en los reactivos, el equilibrio tiende a desplazarse hacia los productos. Calor y Trabajo: NO SON FUNCIONES DE ESTADO El intercambiode energíaentre el sistemaysusalrededoresocurre enformade calor (q) y trabajo (w). Estas no son funciones de estado ya que a diferencia de estas; q y w solo se presentan cuandoocurre un cambioenel sistema y sus valores dependerán del tipo de transformación que experimenta el sistema al pasar de su estado inicial a un estado final. Leyes de la Termodinámica 1a Ley de la Termodinámica (Ley de la Conservación de la Energía) Durante cualquier proceso la energía total del universo se mantiene constante. La energía no puede ser creada o destruida pero si transformada.
- 2. Representación matemática ΔE = Q - w Donde: ΔE = Cambioenla energíainternadel sistema(Joules,J). Q= Calor intercambiadoporel sistemaconel entorno (Joules,J). W= Trabajo intercambiadoporel sistemaconel entorno (Joules,J). Tambiénse puede expresarcomo: ΔE = Ef - Ei Donde ΔE = Cambioenla energíainternadel sistema. Ef = Energía internadel sistemaensuestadoinicial. Ei = Energía internadel sistemaensuestadofinal 2a Ley de la Termodinámica (Ley de la espontaneidad termodinámica) El universosiempre tenderáal incrementode suentropía(medidade desordeno aleatoriedad). Durante cualquierprocesode cambio yase físicoo químico,loselementosdel universotenderána desordenarse. Expresión matemática En un proceso espontáneo: ΔSsistema + ΔSentorno = ΔSuniverso > 0 En un proceso en equilibrio: ΔSsistema + ΔSentorno = ΔSuniverso = 0 Energía libre de Gibbs La energía libre de Gibbs se define como la cantidad de energía capaz de realizar un trabajo durante una reacción a condiciones constantes. Puede ser usada como un indicador de la espontaneidad de una reacción. Gibbs y su relación con la constante de equilibrio (Keq) La constante de equilibrio de una reacción, es la expresión de la proporción que hay entre la concentración de reactantes y la concentración de productos en el punto de equilibrio de la reacción. Keq = [𝑃𝑟𝑜𝑑𝑢𝑐𝑡𝑜𝑠] 𝑒𝑞 [𝑅𝑒𝑎𝑐𝑡𝑖 𝑣 𝑜𝑠] 𝑒𝑞 El ΔG permite calcular que tan alejada se encuentra la reacción del equilibrio bajo condiciones específicas y que tanta energía se liberara conforme la reacción proceda hacia al equilibrio, por ejemplo: En la reacción A B para calcularΔG usandola concentracione prevalentede reactantes(A) y productos (B) se utiliza la ecuación: ΔG=RTln [𝐵] 𝑝𝑟 [𝐴] 𝑝𝑟 - RTln [𝐵] 𝑒𝑞 [𝐴] 𝑒𝑞 = RTln [𝐵] 𝑝𝑟 [𝐴] 𝑝𝑟 – RTln Keq Donde: ΔG= Cambio de energía libre (cal/mol). R=Constante de los gases ideales (1.987cal/mol.K). T=Temperatura (K). [𝐴] 𝑝𝑟= Concentración prevalente de reactivos (moles/litro, M).
- 3. [𝐵] 𝑝𝑟:= Concentración prevalente de productos (moles/litro, M). Gráfico de relación: Energía libre y Equilibrio químico Se muestra que la cantidad de energía libre disponible para una reacción depende de que tan alejados estén los componentes de la reacción del equilibrio. La energía libre del sistema incrementa conforme la proporción de [ 𝐵]/[ 𝐴] cambia a cada lado del punto de equilibrio Otra formade mostrar larelaciónde la energíalibre yla constante de equilibrio es; calculando el cambio de energía libre estándar (ΔG0 ´) de una reacción utilizando el valor de su Keq,. Para ello se puede utilizar la siguiente ecuación: ΔG0 ´= -2.3RTlog Keq=-1362log Keq También se puede calcular Keq a patir de ΔG0 ´ si se reacomoda la ecuación anterior y se toma el antilogaritmo: Keq=10−(ΔG0´/2.3RT) A partirde estaexpresiónse puede deducirque: Si ΔG0 ´ en negativo,el exponente serápositivoyportantoel valorde Keq serámayor a 1. Se favorecerálaformaciónde productos. Si ΔG0 ´ en positivo,el exponenteseránegativoyportantoel valorde Keq serámenora 1. Se favorecerálaformación de reactivos. Otra forma se representar la primera ecuación mostrada y calcular ΔG0 ´ a partir de su Keq en condiciones estándar es: ΔG0 ´= -RTlnK´eq = -RTlnK´eq+ RTln1 = -RTlnK´eq (* El factor 2.3 de la ecuación original permita convertir entre el uso de logaritmo natural (ln) y el logaritmo base 10 (log10).) Gráfico de relación de Energía libre estándar (ΔG0 ´) vs Constante de Equilibrio (K´eq) Se observa que cuando el valor de la constante de equilibrio es 1, el cambio de energíalibre estándares0. Esto se debe a que al sustituir el valor de Keq en la ecuación, al calcular el logaritmo natural de 1 este resulta 0 y por tanto el valor final de ΔG0 ´ también será 0. Tambiénse observa como a valores menores de 1 para la constante de equilibrio, los valores de energía libre estándar son positivosyenel caso de valores mayores de 1 para la constante de equilibrio los valores de energía libre estándar son negativos.
- 4. Relación de los valores de ΔG0 ´con K´eq en una reacción a. ΔG0 ´ NEGATIVO y K´eq >1 En condiciones estándar la reacción es espontánea en dirección a la formación de productos (hacia la derecha). La concentración de productos predomina por sobre la concentración de reactivos. b. ΔG0 ´ POSITIVO y K´eq < 1 En condiciones estándar la reacción es espontánea en dirección a la formación de reactantes (hacia la izquierda). La concentración de reactantes predomina por sobre la concentración de productos. c. ΔG0 ´= 0 y K´eq =1 En condiciones estándar la reacción se encuentra en equilibrio. La concentración de reactivos y productos es igual. Principio de Le Chatelier “Cuandoen unsistemaenequilibrio se produce la modificación de al menos una de las variables que lo determinan (funciones de estado) el equilibrio se desplazará en el sentido en el que se opone a dicha variación” Cualquier desviación del equilibrio del sistema estimula un proceso que tiende a restaurar el equilibrio del sistema. En sistemas cerrados Si un sistema es del tipo cerrado (no hay intercambio de materia con el resto del universo, y sus alrededores) cualquierpequeñocambiode energíainternadel sistema es debido al balance de la transferencia de calor y el trabajo realizado por o sobre el sistema. Cualquier cambio y la recuperación del estado de equilibrio dentro del sistema se basarán principalmente en el intercambio de energía en forma de calor y trabajo. En sistemas vivos Los sistemas vivos son sistemas de tipo abierto; por lo que se encuentran en constante intercambio tanto de materia como de energía con sus alrededores. El comportamiento de la cinética de las reacciones y procesos que se realizan dentro de los sistemas vivos está en constante cambio debido a las variaciones de los requerimientos que se presente enmomentos determinados ya laexposición adistintos factores delentorno.Cualquiera que seael caso, lossistemasvivos tenderánaalejarse cadavez más del equilibrio que mantienen bajo condiciones fisiológicas. Una vez que pase en efecto inmediato del factor que causo la perturbación el sistematenderáadirigirse aunadirección que permitacontrarrestar el efecto del disturbio. Larecuperación del equilibrio se relazará a través del intercambio y transformación de materia y energía. Relación de energía libre de Gibbs, la constante de equilibrio (Keq) y el potencial de reducción El potencial de reducción estándar de los sistemas de oxido-reducción permite predecir la direccióndel flujode electronesde unsistemaaotro,bajo condicionesestándar. Portantoal igual
- 5. que la energía libre de Gibbs el potencial de reducción (E) también puede ser usado para determinar la espontaneidad de una reacción. Tomando como guías, se utilizarán ecuaciones que las relacionan (en condiciones estándar): Relación ΔG0 ´ y Keq ΔG0 ´= -RTlnK´eq Donde: ΔG= Cambio de energía libre (cal/mol). R= Constante de los gases ideales (1.987cal/mol.K). T= Temperatura (K). K´eq=constante de equilibrio de la reacción. Relación ΔG0 ´ y E0 ´ Ecuación de Nernst ΔG0 ´= -nFΔE0 ´ Donde: ΔG0 = Cambiode energíalibre estándar(cal/mol). n= Númerode electronestransferidosenlareacciónredox. F= Constante de Faraday (23.060cal/voltioso96.487coulomb/mol) ΔE0 ´= Diferenciaenel potencialde reducción (Volts,V). *El signonegativo(-) enla fórmulaindicaque se pierde energía, pararealizaruntrabajo. La ecuación de Nernst permite calcular el cambio de energía libre estándar de una reacción de oxido-reducción a partir de la diferencia de los potenciales redox de un par redox. Un ΔE0 ´ con valor positivo, indica que la reacción es espontánea. Una diferencia positiva en el cambio de potencial de reducción conllevara a un cambio negativo de la energía libre estándar. +ΔE0 ´ -ΔG0´ Relación E0 ´ y Keq E0 ´= 𝑅𝑇 𝑛𝐹 𝑙𝑛 [𝐹𝑜𝑟𝑚𝑎 𝑂𝑥𝑖 𝑑 𝑎𝑑𝑎] [𝐹𝑜𝑟𝑚𝑎 𝑅𝑒𝑑𝑢𝑐𝑖 𝑑 𝑎] E0 ´= 𝑅𝑇 𝑛𝐹 𝑙𝑛Keq Donde: ΔE0 ´: Diferenciaenel potencial de reducción (Voltios). R: Constante de los gases ideales (1.987cal/mol.K). T: Temperatura (K). Keq: constante de equilibrio de la reacción (Calculada con las concentraciones de las formas oxidadas y reducidas del par redox de la reacción). n:númerode electronestransferidosenla reacciónredox. F: Constante de Faraday(23.060cal/voltioso 96.487coulomb/mol) Procesos Espontáneos Algunos parámetros termodinámicos útiles para predecir la espontaneidad de un proceso son:
- 6. Energía libre de Gibbs Entropía Entalpía Potencial Reducción Procesos en Sistemas Vivos 1.-Ejemplosdeprocesosespontáneos 1.1) Trasporte de sustancias a través de la membrana por gradiente de concentración Soloocurrirá uncambioen el sistemasi hay un gradiente de concentración. La dirección que tomen las partículas dependerá de las concentraciones encontradas en ambos lados de la membrana, ya que el potencial o contenido químico es el mismo a ser la misma molécula. 1.2) Hidrólisis de ATP La hidrólisisdel ATPesunprocesoespontáneo, que permite acoplarestareacciónexergónicaa procesos o reacciones endergónicas, es decir, que necesitan un aporte energético 2.-Ejemplosdeprocesosno espontáneos Los procesos de la célula que por naturaleza serían termodinámicamente no espontáneos, se encuentran acoplados a otros procesos o elementos que vuelven factible su realización. 2.1) Plegamientode proteínas El plegamiento de proteínas se considera un proceso por naturaleza termodinámicamente no espontáneo. Pero en condiciones celulares cuenta con la ayuda de enzimas, azúcares, y otros elementos que se acoplan a este para que pueda desarrollarse. 3.- Ejemplosde procesosacoplados 3.1) Acoplamientode reacciones Un ejemplo es el primer paso de la glucolisis; el cambio de la glucosa o glucosa-6-fosfato: La reaccióndirectanoes favorable termodinámicamente ya que su ΔG0 ´ es positiva, para volverla favorables es acoplada a la hidrólisis de ATP. Esto hace que los valores acoplados de ΔG0 ´ de ambas reaccionesdenunvalornegativoyportanto el procesose termodinámicamentefavorable. Glucosa + Pi Glucosa-6-P ΔG0 ´=+13.8kJ/mol ATP ADP + Pi ΔG0 ´=-30.5kJ/mol Glucosa+ ATP Glucosa-6-P+ ADP ΔG0 ´ =-16.5 kJ/mol 3.2) Acoplamientode procesos:Anabolismoy Catabolismo
- 7. Un ejemplomásgeneral de procesosacoplados, es la transferencia de energía del catabolismo al anabolismo, que se denomina acoplamiento energético. Anabolismo y catabolismo están acoplados, de forma que la energía y el poder reductor que generanlasreaccionesdel catabolismo son consumidas por las del anabolismo. Pero como estos procesosnoocurren ni de forma simultánea, ni en el mismo lugar de la célula, se necesitan unas sustancias:intermediarios del metabolismo,que transporten la energía, los electrones y el poder reductordesde donde se generanhastadonde se necesitan. Estos intermediarios son el ATP y las coenzimas transportadores de electrones. Ejemplos de Sistemas Aislados El sistemaaisladoidealesteóricoporque requierede unafronterafísicaque seaun aislante yeste no existe. Algunos ejemplos aproximados son: o Un termo: Durante un período de tiempo, los termos logran aislar el contenido en su interioryevitarlafuga de energíahacia el medioambiente,yal mismotiempoimpiden el derramamiento del contenido o la introducción del mismo. o El universo:nose sabe qué hayafuerade él,ni si hay un afuera de él, por lo que se podría suponer que el universo es un sistema aislado, en el que no ingresa ni egresa materia o energía, opera de manera independiente de su entorno. o Productos enlatados: En condiciones normales, estos productos se encuentran lejos de cualquier intercambio de materia o de energía, a menos que sean expuestos a alguna condición que provoque cambio de temperatura, etc. o Trajes de neopreno: estos no permiten el intercambio de energía entre el agua y el nadador. o Cámaras hiperbáricas: sonutilizadasparaaislarde lascondicionesatmosféricasalosbuzos con burbujas de nitrógeno en la sangre, no permiten el intercambio de materia ni de energía, o al menos no en cantidades apreciables y significativas. Hidrólisis de ATP La energíaútil de lamoléculade ATPse encuentracontenida en sus enlaces fosfoanhídridos. Esta energía está disponible para su uso cuando uno de estos enlaces es hidrolizado. La hidrólisisde estosenlacespuede ocurrir debido a la repulsión que existe entre los átomos de oxígenocargadosnegativamenteque se encuentranenlamoléculade ATP.Estarepulsión provoca que la estructura del ATP sea muy inestable y por lo tanto, se tenderá a liberar esta tensión. Cada enlace fosfoanhídrido tiene una ΔG0 ´ de aproximadamente -7.3kcal/mol por hidrólisis. El ATPes el intermediario común en la mayoría de las reacciones no espontáneas de la célula, ya que la transferenciadel grupo fosforilo por hidrólisis de ATP es termodinámicamente favorable. Cuando una reacción no es energéticamente favorable esta puede ser acoplada a otra reacción muy favorable (en este caso la hidrólisis de ATP).
- 8. La energía liberada por la hidrólisis del enlace fosfoanhídrido será transferida a uno de los reactantes (B) de la reacción no favorable. El grupo fosfato (Pi) liberado del ATP se une a covalentemente a este reactante (B), generando un intermediario que puede reaccionar con el segundo reactante (C), y se lleva finalmente a la producción de productos y Pi. La reaccióngeneral sería: Planteamiento de un Problema La siguiente reacción reversible es un paso en la vía de la glucolisis: 3-FOSFOGLICERATO 2-FOSFOGLICERATO ---------------------------------(1) (3PG) (2PG) Cuando se adiciona la enzima que cataliza esta reacción a una solución acuosa de 3PG a 25°C y pH de 7, el valor de la constante de equilibrio es de 0.165. Las concentraciones prevalente de estas substancias en los glóbulos rojos de humano son: 61 micromolar para 3PG y 4.3 micromolar para 2PG. Calcular: a. ΔG0 ´. Explicar con sus propias palabras el significado del valor obtenido ΔG0 ´= -RTlnKeq ΔG0 ´ = -(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(ln0.165)= -(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(-1.802) ΔG0 ´=1067.011𝑐𝑎𝑙 𝑚𝑜𝑙 Es una reacción no espontánea bajo condiciones estándar (donde los reactantes y productos se encuentran en concentraciones iniciales de 1M) al alcanzar la reacción su punto de equilibrio, el 3-Fosofoglicerato predominarían por sobre el 2-Fosfoglicerato es decir, la reacción tenderá a la izquierda. b. CalcularΔG´.Explicarqué significadotiene estevalory porqué ΔG´ y ΔG0 ´ son diferentes. ΔG´= RTln[𝑃𝑟𝑜𝑑𝑢𝑐𝑡𝑜𝑠] [𝑅𝑒𝑎𝑐𝑡𝑎𝑛𝑡𝑒𝑠] - RTlnKeq ΔG´= RTln[2𝑃𝐺] [3𝑃𝐺] - RTlnKeq ΔG´= (1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(ln (4.3𝑋10 −6 𝑀) (61𝑋10−6 𝑀) ) -(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙 .𝐾 )(298K)(ln0.165) ΔG´= (1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(ln 0.070) -(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙 .𝐾 )(298K)(ln0.165) ΔG´= (1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(-2.659) -(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(-1.802) ΔG´=-1574.463 𝑐𝑎𝑙 𝑚𝑜𝑙 +1067.011 𝑐𝑎𝑙 𝑚𝑜𝑙 ΔG´=-507.452 𝑐𝑎𝑙 𝑚𝑜𝑙 Bajo las condiciones prevalentes en la célula, la reacción es termodinámicamente favorable, por tanto tiende a la derecha (formación de productos). Los valoresde ΔG´ y ΔG0 ´ sondistintosdebidoa que al calcular ΔG0 ´ se utilizan valores de condicionesestándar,enestascondicionesse establece que las concentraciones iniciales tanto de 3-fosfoglicerato como de 6-fosfoglicerato son de 1M. En el caso del cálculo de ΔG´ se toman en cuenta los valores prevalentes (en condiciones celulares) de lasconcentracionesde losreactivosy los productos, que en esta caso son 61 µM para 3PG y 4.3 µM para 2PG.
- 9. c. Considerandoque laconcentraciónde 3PG permanece constante en 61 micromolar, pero la concentración de 2PG se incrementa, calcular que tanto puede elevarse la concentración de 2PG antes de que la reacción se detenga porque ya no es termodinámicamente factible. ΔG´− RTlnKeq 𝑅𝑇 = ln[2𝑃𝐺] [3𝑃𝐺] Para quela reacción llegue al equilibrio: 0+(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(ln0.165) (1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K) = ln [2𝑃𝐺] (61𝑋 10−6 𝑀) 0+(1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K)(−1.802) (1.987 𝑐𝑎𝑙 𝑚𝑜𝑙.𝐾 )(298K) = ln [2𝑃𝐺] (61𝑋10−6 𝑀) −1067.011 𝑐𝑎𝑙 𝑚𝑜𝑙 592.126 𝑐𝑎𝑙 𝑚𝑜𝑙 = ln [2𝑃𝐺] ( 61𝑋10−6 𝑀) −1.802 = ln [2𝑃𝐺] (61𝑋10−6 𝑀) e-1.802 = [2𝑃𝐺] (61𝑋 10−6 𝑀) (0.165)(61x10-6 M)= [2𝑃𝐺] [2PG]=1.007X10-5 M Para que la reacciónllegue al puntode equilibrio se requeriría aumentar la concentración de 2-Fosfoglicerato de 4.3X10-6 Ma 1.007X10-5 M(diferencia de 5.770X10-6 M). Si se supera la concentración de 2PG más allá de este punto la reacción se volverá termodinámicamente no favorable.