Descargado 727 veces



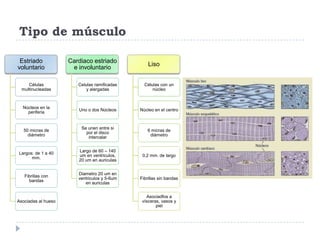
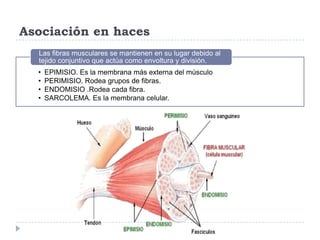



























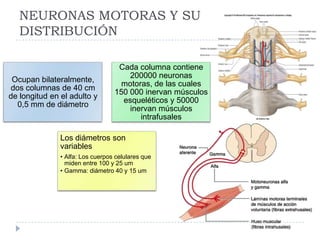



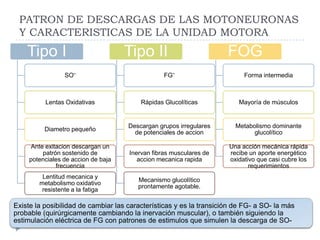










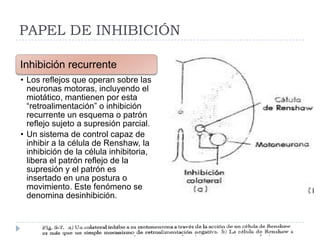





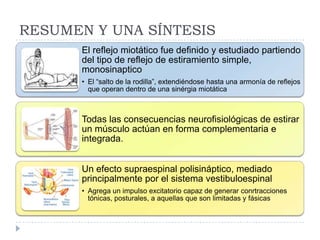









El documento describe la estructura y tipos de músculo, incluyendo el músculo estriado esquelético, cardiaco y liso. También explica el reflejo miotático, el cual ocurre cuando los receptores en los husos musculares detectan un estiramiento muscular y envían señales a la médula espinal para provocar una contracción y evitar la elongación. Finalmente, detalla las neuronas motoras en la médula espinal y su papel en el reclutamiento muscular según el principio de tamaño de Henneman.























































































