Descargado 71 veces

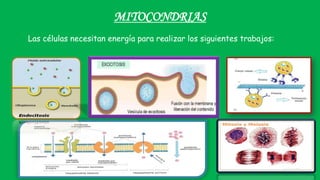

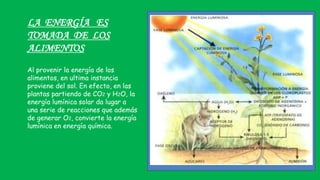


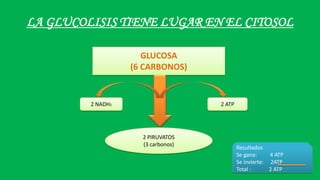
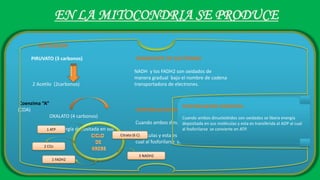
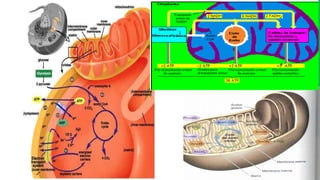


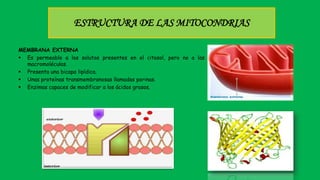

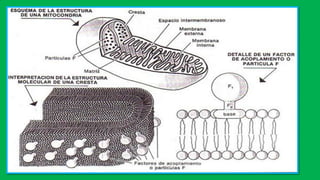

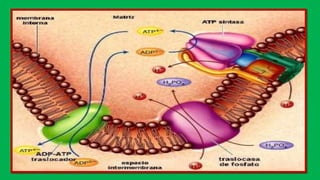
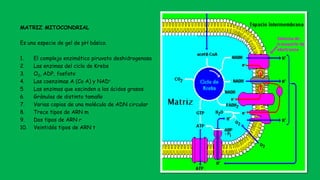


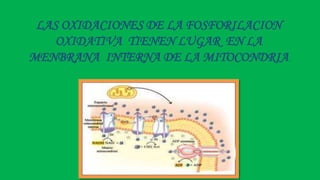


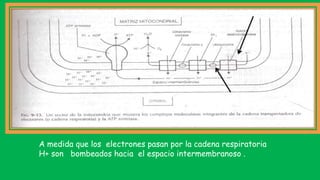
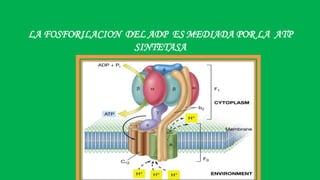
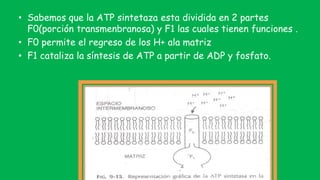


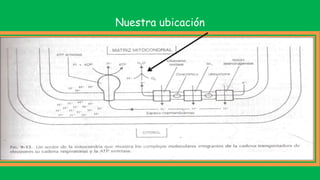



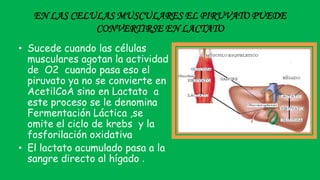





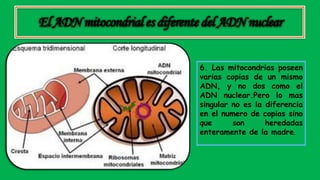


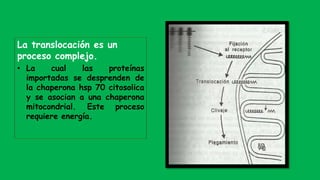







El documento describe las funciones de las mitocondrias en las células. Las mitocondrias generan la mayor parte de la energía de las células a través de la fosforilación oxidativa, donde la energía de los alimentos se convierte en ATP. Las mitocondrias contienen cadenas de transporte de electrones en su membrana interna que producen ATP a medida que los electrones pasan a través de ellas. También albergan varios ciclos metabólicos como la glucólisis y el ciclo de Krebs.











































































































