Descargado 642 veces





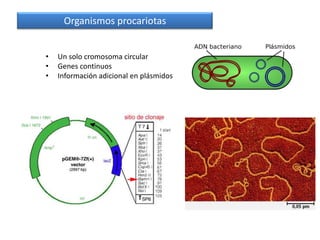




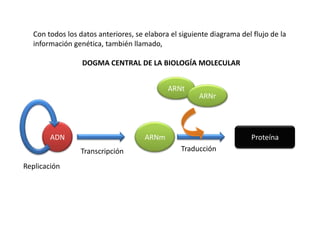
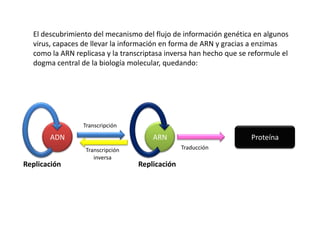



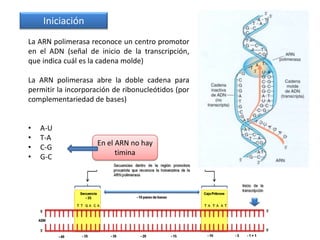



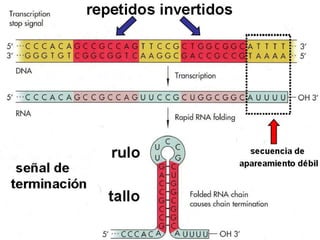

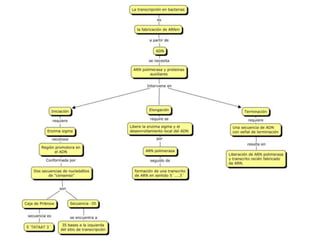
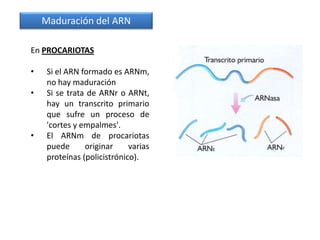










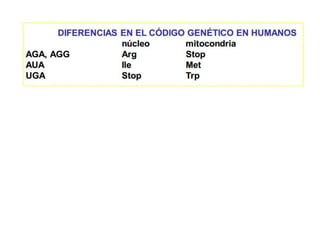
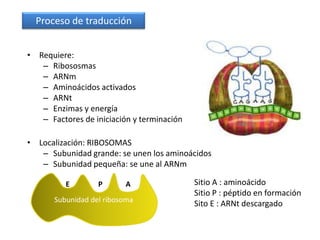


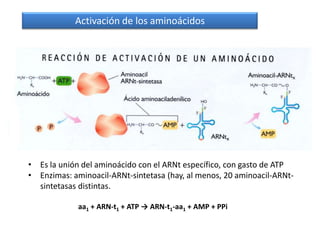

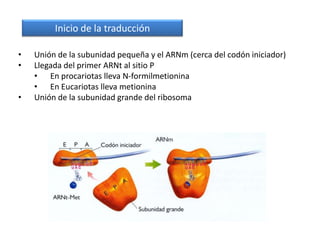



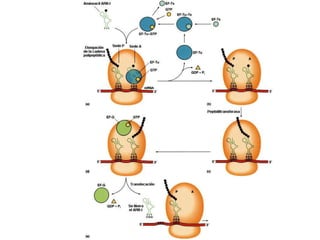
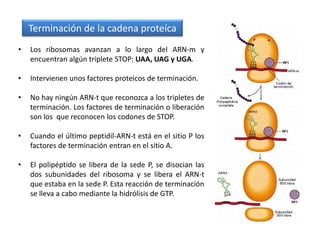
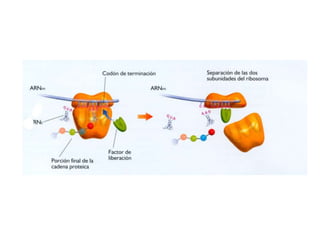

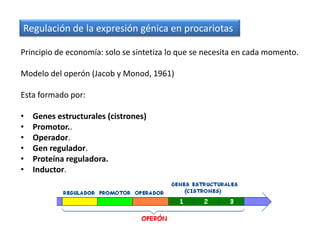

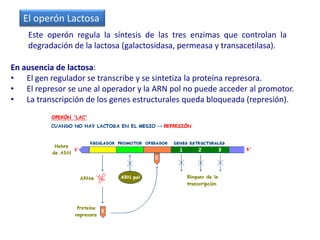



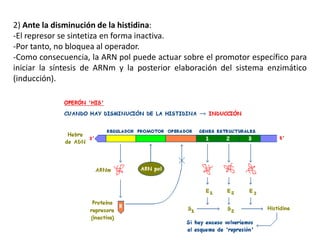




El documento describe el descubrimiento del flujo de la información genética desde el ADN hasta la síntesis de proteínas. Se detalla el proceso de transcripción del ADN en ARNm, y la traducción del ARNm en proteínas por los ribosomas. El código genético fue descifrado experimentalmente, determinándose que está formado por tripletes de nucleótidos (codones) que especifican los aminoácidos.




























































![GENETICA NOVENO GRADO [Autoguardado].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/geneticanovenogradoautoguardado-250914145105-0dc0f10e-thumbnail.jpg?width=640&height=640&fit=bounds)














































