Descargado 138 veces
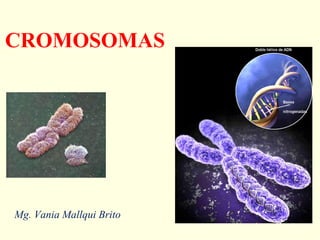
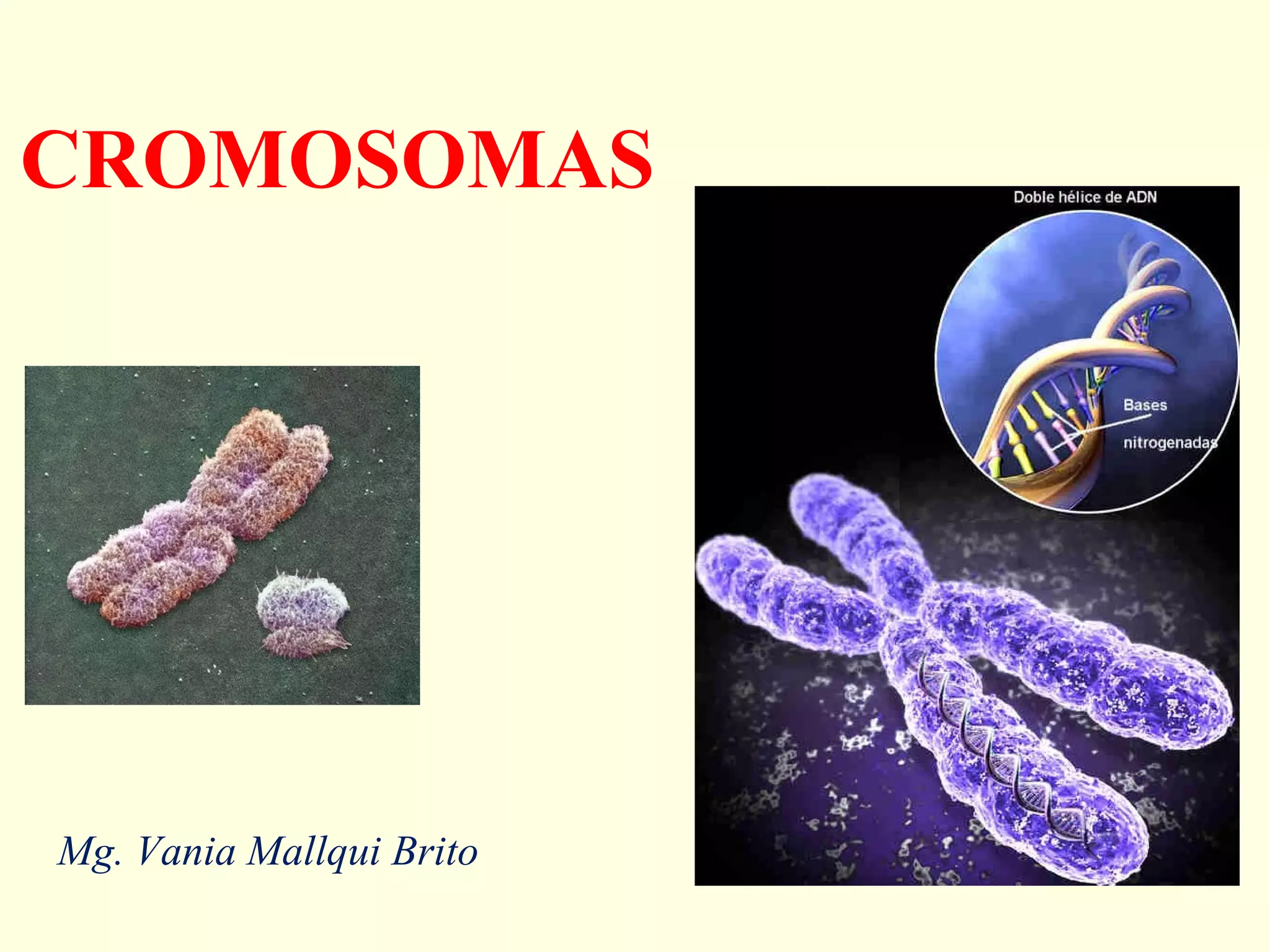



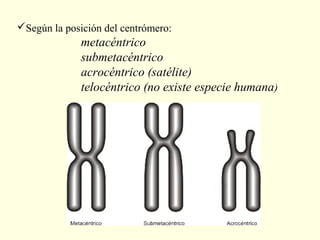

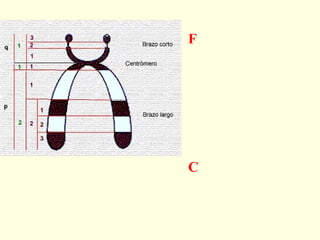
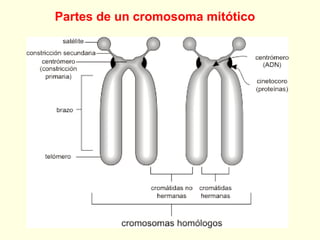
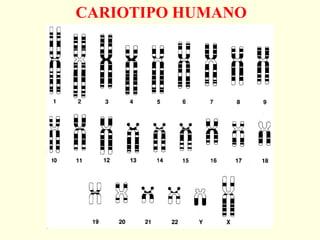

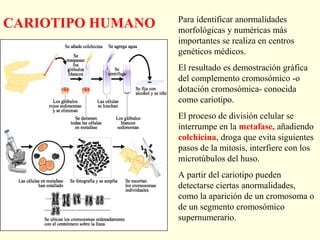


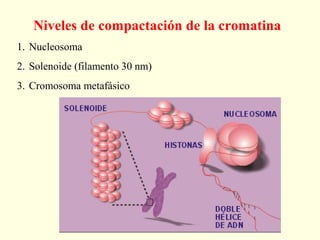

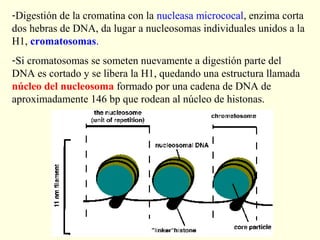
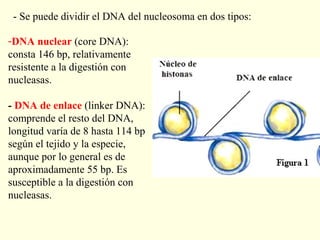


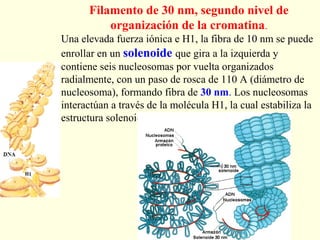
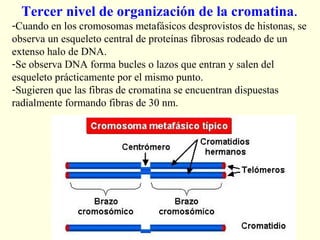
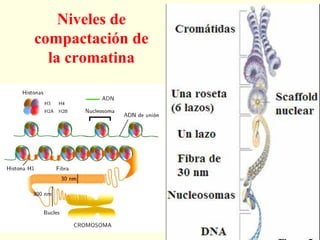

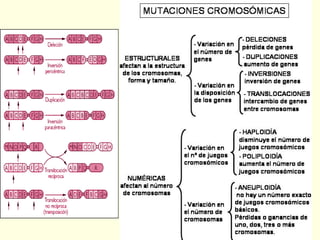




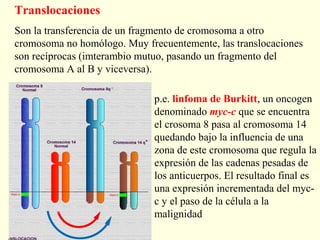
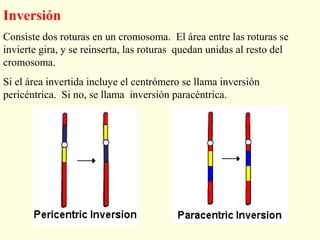

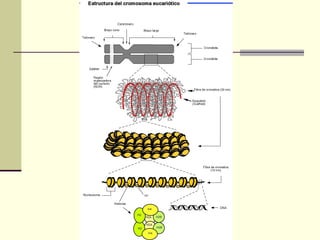

El documento describe los cromosomas, que son portadores del material genético y determinan las características hereditarias de cada especie. Explica que los cromosomas se pueden ver al microscopio durante la metafase y contienen ADN empaquetado en cromátidas unidas por un centrómero. También describe las partes de los cromosomas como los telómeros, centrómeros y cinetócoros, y explica cómo los cromosomas se organizan en el cariotipo humano.

























![Transcripsion en eucariotas[1]](https://cdn.slidesharecdn.com/ss_thumbnails/transcripsioneneucariotas1-121021194324-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)






















































